| 内容要旨 | | センダイウイルス(Hemagglutinating virus of Japan,以下HVJと略す)は,パラミクソウイルス科に属し,約15,000ヌクレオチドからなる非分節マイナス鎖RNAをゲノムに持つ。ゲノムは3’側よりNP,P/C,M,F,HN,Lの順にタンパク質をコードし,3’および5’末端には,それぞれ約50ヌクレオチドから成るタンパク質をコードしないリーダー領域が存在する。類似したゲノム構造を持つウイルスとしては,同じ科に属する麻疹ウイルス,ムンプスウイルスなどが,またラブドウイルス科に属するウシ水疱性口内炎ウイルス,狂犬病ウイルスなどが挙げられる。感染細胞内で,ウイルスゲノムの持つ遺伝情報は6種類のモノシストロニックmRNAを介して発現される。これらのウイルスmRNAは,5’末端にキャップ構造,3’末端にポリ(A)鎖を有する典型的な真核細胞型のmRNAの構造を持ち,宿主のタンパク質生合成装置を利用して翻訳される。一方,ゲノムの複製はゲノム全長に対応する完全鎖長のプラス鎖RNA中間体を経て行なわれる。転写,複製反応ともにウイルス粒子に含まれるRNA依存RNAポリメレースが触媒すると考えられている。 我々は,HVJゲノムの転写,複製の分子機構の解明を目的に,精製ウイルス粒子を用いたin vitro転写系を確立し,その解析を行なって,これまでに以下の結果を得てきた。(1)HVJの転写反応には,真核細胞に普遍的に存在する宿主由来のタンパク質性因子(宿主因子)が必須である。(2)in vitro転写系で合成される転写産物は,その多くが,感染細胞内で合成されるウイルスmRNAと同様,5’キャップ構造(m7GpppAm),3’ポリ(A)鎖を有しており,また,ゲノム上の正確な領域から転写されたプラス鎖である。(3)高い宿主因子活性を有するウシ脳抽出液より因子の部分精製を試み,単独では低い活性しか示さないが,組み合わせると相乗的な活性を示す二つの相補的な画分に分かれることを見出した。その一つは,高度に精製したチューブリンで置き換えることが可能であったことから,宿主因子は少なくとも二種の成分からなり,その一つはチューブリンであると推定された。 本研究では,HVJの転写に必須な宿主因子を同定し,その機能を解明することを目的として,[1]部分精製宿主因子標品の性質,[2]転写開始複合体の形成反応,[3]宿主因子活性の動物臓器分布,の三項目について種々の解析を行なった。 [1]ウシ脳抽出液より,ハイドロキシルアパタイトカラムクロマトグラフィーにより部分精製される二つの相補的な画分について解析した。まずイムノブロット解析を行ない,一方の画分にのみチューブリンが検出されることを見出した。チューブリンが検出されない画分に含まれる因子は,ゲル濾過法による分析で分子量約40,000と推定された。チューブリンを含んでいる画分は,チューブリンのみならずポリグルタミン酸やポリアスパラギン酸でも置き換えが可能であったことから,宿主因子としてのチューブリンの機能ドメインはC末端近傍の酸性アミノ酸残基に富む領域である可能性が考えられた。 [2]これまでに解析を行なってきた,ウイルス粒子を用いた一段階のインキュベーションによるoverallのRNA合成反応を,開始段階と伸長段階以後,という二つの素過程に分けて解析した。特に開始段階における宿主因子の機能を明らかにすることを目的に,転写開始複合体の単離を試みた。ウイルス粒子とウシ脳抽出液とを基質ヌクレオチド(4NTPs)非存在下にプレインキユベートした後,ヌクレオカプシド(RNP)画分を回収した。この画分には4NTPsのみの添加によりoverall反応系における転写産物に等しい大きさのRNAを合成する活性が検出されたことから,プレインキュベーションにより転写開始複合体が形成されてRNPとして単離されたことが示された。ウイルスの各遺伝子に対するcDNAを用いたスロットブロット解析の結果,このRNA産物には6種類のウイルス遺伝子全てに対応したRNAが含まれていることが示された。さらに5’および3’末端の構造を分析して,このRNAには3’ポリ(A)鎖が付加されており,5’末端についてはm7GpppAmというキャップ構造が存在することを見出した。これらの結果から,転写開始複合体によって合成されるRNA鎖は,基本的にはoverall反応と同様に,ゲノムの正確な領域が転写されたmRNAであり,正しいプロセッシングを受けていることが明らかとなった。またこのことは,これらRNA修飾に関与する酵素活性がRNPを構成するタンパク質のいずれかに存在することを示唆している。 次に,イムノブロット法で転写開始複合体を解析したところ,複合体にはRNPの構成タンパク質以外に,活性の強さにほぼ比例した量の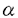 および および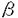 チューブリンが含まれていた。以上の結果から,宿主因子の機能の一つは,RNPに作用して転写開始複合体を形成すること,因子の一つチューブリンは複合体形成反応において,おそらくヘテロダイマーとして複合体に組み込まれて機能していること,の二点が示唆された。 チューブリンが含まれていた。以上の結果から,宿主因子の機能の一つは,RNPに作用して転写開始複合体を形成すること,因子の一つチューブリンは複合体形成反応において,おそらくヘテロダイマーとして複合体に組み込まれて機能していること,の二点が示唆された。 そこで,ウシ脳より高度に精製したチューブリンを用いて転写開始反応を行なったところ,チューブリン単独で開始複合体形成を促進することが明らかとなった。またこの場合にも,転写産物は5’キャップ構造と3’ポリ(A)鎖を有していた。さらに,チューブリンの代わりに[1]に記した合成アミノ酸ポリマーを用いても,かなりの程度置き換えが可能であった。すなわち,チューブリンのみでも,そのC末端近傍の酸性アミノ酸領域が機能することによって正確な転写を行ない得る開始複合体が形成される可能性が示唆された。 次に,先に記した,ハイドロキシルアパタイトカラムにより部分精製された二つの画分を用いて転写開始複合体形成を調べたところ,各画分単独で活性複合体が形成されることが明らかとなった。またこの場合,二つの画分の共存下に転写開始反応を行なっても開始複合体形成に対する相乗効果は認められなかった。イムノブロット解析の結果,overall反応においてチューブリンで置換可能である画分のみによる複合体と,二つの画分を共存させて得られた複合体の二種についてはチューブリンが検出されたが,チューブリンを含んでいない画分のみにより形成された複合体にはチューブリンは検出されなかった。このことから,開始複合体の形成においてチューブリンは必ずしも必須なタンパク質分子ではないと推定される。overall反応で観察された相補性が複合体形成反応においては見られなかったという点については,この相補性が,1)複合体の形成効率に関係している,2)ポリメレースや宿主因子の再利用といった伸長段階以後の素反応におけるものである,等の可能性が考えられる。また,チューブリンを含まない画分を用いても活性複合体が形成された,という点については,この画分中に,開始複合体形成反応においてある程度チューブリンの代役となりうるタンパク質が存在する可能性も考えられる。今後は,チューブリン以外の因子を同定すべくその精製を進めると同時に,反応の素過程における機能を解明していく必要があろう。 [3]HVJはマウスに感染して重篤な肺炎を惹起する。この臓器特異性は気管支上皮に偏在するプロテアーゼにより規定されていることが明らかとされているが,我々の見出した宿主転写因子活性が何らかの影響を及ぼしている可能性を考慮して,ラット各臓器抽出液のoverall転写反応における宿主因子活性を測定した。その結果,HVJ感染HeLa細胞ならびにウシ脳の抽出液に匹敵する高い活性が脳,肺,精巣,卵巣各抽出液に検出され,胸腺,心,肝,脾,子宮を用いても有意なRNA合成が認められた。しかし腎抽出液では活性は検出されなかった。次に各抽出液に内在するRNA分解活性(RNase活性)を調べる目的で,HeLa細胞抽出液を用いたoverall反応でアイソトープ標識したウイルスmRNAを合成し,各抽出液とインキュベートして分解されるRNAの割合を調べた。その結果,RNA合成が全く認められなかった腎には高いRNase活性が検出されたのをはじめ,肝以外の臓器に関しては,宿主因子活性とRNase活性とは負の相関を示す傾向が見られた。すなわち,臓器間の宿主因子活性の差は,肝を除いて,主に抽出液中のRNase活性の差に由来したものであると言える。 そこで,RNase活性の影響を除外することを目的に,各抽出液を用いて転写開始複合体を単離し,RNA合成活性を検討したところ,上記overall反応で活性が極めて低い,あるいは全く検出されなかった肝,腎を含めて,いずれの抽出液を用いても活性複合体が形成されることが示された。このことから,検討した臓器全てに少なくとも開始複合体形成に必要な宿主因子が存在することが明らかとなり,また,overall反応における宿主因子活性の差が主にRNase活性の差に基づくものであるという推定が裏付けられた。これらの複合体をイムノブロット法で解析したところ,全てにチューブリンが検出された。以上の結果は,宿主因子の一つがチューブリンであるという推測を支持するものであり,また,宿主転写因子活性が臓器特異性を大きく規定するものではないことを示唆している。 しかしながら,これらの解析において,肝抽出液はRNase活性がほとんど検出されないにも拘らずoverall転写反応における活性が著しく低いことが見出された。この点に着目して,HeLa細胞抽出液によって促進されるoverall転写反応に対する各臓器抽出液の影響を検討したところ,肝抽出液にのみ転写の全体的な抑制効果が認められ,HVJの転写を抑制する因子の存在が示唆された。この抑制因子活性はゲル濾過法による分析で分子量約100,000と推定された。また,活性画分はRNase活性を全く示さなかった。これらの結果は,HVJの転写反応が正の因子のみならず負の因子によっても制御されている可能性を示唆するものである。今後,この転写抑制因子の精製をも試み,肝特異的なものか否かを検討すると同時に,転写の素過程における作用機作を解析することは,HVJ増殖に関与する宿主タンパク質群の役割を理解する上で興味ある問題である。 |
| 審査要旨 | | 本研究は,マイナス鎖RNAゲノムの転写,複製の分子機構の解明を目的に,パラミクソウイルス科に属するセンダイウイルス(Hemagglutinating virus of Japan,以下HVJと略す)の,精製ウイルス粒子を用いたin vitro転写系を駆使して,HVJゲノムの遺伝情報発現機構の解析を行なったものである。特に,in vitro転写反応に必須な宿主由来のタンパク質性因子(宿主因子)に着目し,その同定ならびに機能解析を行ない,以下の結果を得ている。 1.これまでに,申請者の共同研究者が,高い宿主因子活性を有するウシ脳抽出液より因子の部分精製を試み,ハイドロキシルアパタイトカラムクロマトグラフィにより,単独でに低い活性しか示さないが,組み合わせると相乗的な活性を示す二つの相補的な画分に分かれること,その一つは,高度に精製したチューブリンで置き換えることが可能であったことから,宿主因子は少なくとも二種の成分からなり,その一つはチューブリンであると推定されること,を見出していた。申請者は,この二つの相補的な宿主因子部分精製標品について解析した。まずイムノブロット解析を行ない,一方の画分にのみチューブリンが検出されることを見出した。チューブリンが検出されない画分に含まれる因子は,ゲル濾過法による分析で分子量約40,000と推定された。チューブリンを含んでいる画分は,チューブリンのみならずポリグルタミン酸やポリアスパラギン酸でも置き換えが可能であったことから,宿主因子としてのチューブリンの機能ドメインはC末端近傍の酸性アミノ酸残基に富む領域である可能性が考えられた。 2.これまでに解析を行なってきた,ウイルス粒子を用いた一段階のインキュベーションによるoverallのRNA合成反応を,開始段階と伸長段階以後,という二つの素過程に分けて解析した。特に開始段階における宿主因子の機能を明らかにすることを目的に,転写開始複合体の単離を試みた。ウイルス粒子とウシ脳抽出液とを基質ヌクレオチド(4NTPs)非存在下にプレインキユベートした後,ヌクレオカプシド(RNP)画分を回収した。この画分には4NTPsのみの添加によりoverall反応系における転写産物に等しい大きさのRNAを合成する活性が検出されたことから,プレインキュベーションにより転写開始複合体が形成されてRNPとして単離されたことが示された。ウイルスの各遺伝子に対するcDNAを用いたスロットブロット解析の結果,このRNA産物には6種類のウィルス遺伝子全てに対応したRNAが含まれていることが示された。さらに5’および3’末端の構造を分析して,このRNAには3’ポリ(A)鎖が付加されており,5’末端についてはm7GpppAmというキャップ構造が存在することを見出した。これらの結果から,転写開始複合体によって合成されるRNA鎖は,基本的にはoverall反応と同様に,ゲノムの正確な領域が転写されたmRNAであり,正しいプロセッシングを受けていることが明らかとなった。またこのことは,これらRNA修飾に関与する酵素活性がRNPを構成するタンパク質のいずれかに存在することを示唆している。 次に,イムノブロット法で転写開始複合体を解析したところ,複合体にはRNPの構成タンパク質以外に,活性の強さにほぼ比例した量のおよびチューブリンが含まれていた。以上の結果から,宿主因子の機能の一つは,RNPに作用して転写開始複合体を形成すること,因子の一つチューブリンは複合体形成反応において,おそらくヘテロダイマーとして複合体に組み込まれて機能していること,の二点が示唆された。 そこで,ウシ脳より高度に精製したチューブリンを用いて転写開始反応を行なったところ,チューブリン単独で開始複合体形成を促進することが明らかとなった。またこの場合にも,転写産物は5’キャップ構造と3’ポリ(A)鎖を有しており,また,6種類のウイルス遺伝子全てに対応したRNAが含まれていた。さらに,チューブリンの代わりに1に記した合成アミノ酸ポリマーを用いても,かなりの程度置き換えが可能であった。すなわち,チューブリンのみでも,そのC末端近傍の酸性アミノ酸領域が機能することによって正確な転写を行ない得る開始複合体が形成される可能性が示唆された。 次に,先に記した,ハイドロキシルアパタイトカラムにより部分精製された二つの画分を用いて転写開始複合体形成を調べたところ,各画分単独で活性複合体が形成されることが明らかとなった。またこの場合,二つの画分の共存下に転写開始反応を行なっても開始複合体形成に対する相乗効果は認められなかった。イムノブロット解析の結果,overall反応においてチューブリンで置換可能である画分のみによる複合体と,二つの画分を共存させて得られた複合体の二種についてはチューブリンが検出されたが,チューブリンを含んでいない画分のみにより形成された複合体にはチューブリンは検出されなかった。このことから,開始複合体の形成においてチューブリンは必ずしも必須なタンパク質分子ではないと推定される。overall反応で観察された相補性が複合体形成反応においては見られなかったという点については,この相補性が,1)複合体の形成効率に関係している,2)ポリメレースや宿主因子の再利用といった伸長段階以後の素反応におけるものである,等の可能性が考えられる。また,チューブリンを含まない画分を用いても活性複合体が形成された,という点については,この画分中に,開始複合体形成反応においてある程度チューブリンの代役となりうるタンパク質が存在する可能性も考えられる。以上の点を考慮し,今後,チューブリン以外の因子をも同定し,反応の素過程における機能を解明する必要があることが示された。 [3]HVJはマウスに感染して重篤な肺炎を惹起する。この臓器特異性は気管支上皮に偏在するプロテアーゼにより規定されていることが明らかとされているが,我々の見出した宿主転写因子活性が何らかの影響を及ぼしている可能性を考慮して,ラット各臓器抽出液のoverall転写反応における宿主因子活性を測定した。その結果,HVJ感染HeLa細胞ならびにウシ脳の抽出液に匹敵する高い活性が脳,肺,精巣,卵巣各抽出液に検出され,胸腺,心,肝,脾,子宮を用いても有意なRNA合成が認められた。しかし腎抽出液では活性は検出されなかった。次に各抽出液に内在するRNA分解活性(RNase活性)を調べる目的で,HeLa細胞抽出液を用いたoverall反応でアイソトープ標識したウイルスmRNAを合成し,各抽出液とインキュベートして分解されるRNAの割合を調べた。その結果,RNA合成が全く認められなかった腎には高いRNase活性が検出されたのをはじめ,肝以外の臓器に関しては,宿主因子活性とRNase活性とは負の相関を示す傾向が見られた。すなわち,臓器間の宿主因子活性の差は,肝を除いて,主に抽出液中のRNase活性の差に由来したものであると言える。 そこで,RNase活性の影響を除外することを目的に,各抽出液を用いて転写開始複合体を単離し,RNA合成活性を検討したところ,上記overall反応で活性が極めて低い,あるいは全く検出されなかった肝,腎を含めて,いずれの抽出液を用いても活性複合体が形成されることが示された。このことから,検討した臓器全てに少なくとも開始複合体形成に必要な宿主因子が存在することが明らかとなり,また,overall反応における宿主因子活性の差が主にRNase活性の差に基づくものであるという推定が裏付けられた。これらの複合体をイムノブロット法で解析したところ,全てにチューブリンが検出された。以上の結果は,宿主因子の一つがチューブリンであるという推測を支持するものであり,また,宿主転写因子活性が臓器特異性を大きく規定するものではないことを示唆している。 しかしながら,これらの解析において,肝抽出液はRNase活性がほとんど検出されないにも拘らずoverall転写反応における活性が著しく低いことが見出された。この点に着目して,HeLa細胞抽出液によって促進されるoverall転写反応に対する各臓器抽出液の影響を検討したところ,肝抽出液にのみ転写の全体的な抑制効果が認められ,HVJの転写を抑制する因子の存在が示唆された。この抑制因子活性はゲル濾過法による分析で分子量約100,000と推定された。また,活性画分はRNase活性を全く示さなかった。これらの結果より,HVJの転写反応が正の因子のみならず負の因子によっても制御されている可能性が示唆された。今後,この転写抑制因子の精製をも試み,肝特異的なものか否かを検討すると同時に,転写の素過程における作用機作を解析することは,HVJ増殖に関与する宿主タンパク質群の役割を理解する上で興味ある問題であると考えられる。 以上,本論文は,HVJの転写反応に,細胞骨格タンパク質であるチューブリンを含む複数の宿主因子が関与していることを明らかにした。本研究はこれまでに未知に等しかった,RNAウイルスの転写,複製機構の解明に新たな知見をもたらすものであり,ひいては,ウイルスの病原性の分子機構の解明に重要な貢献をなすことが期待され,よって,学位の授与に値するものと考えられる。 |