固体化学の分野において、対象となる物質の薄膜化は物性測定や応用への発展に対し、きわめて重要なブレークスルーとなっている。薄膜成長に対しては、1分子レベルで均一な結晶性の高い膜の作製が望まれるが、このためには薄膜成長機構の理解が不可欠である。現在まで金属や半導体の系において薄膜成長観察が行われてきたが、基板表面に生成した数原子からなる核や単原子膜などの観察は系が不純物によって汚染されやすいため非常に困難であり、核形成機構や単分子層の成長形態など薄膜成長の初期過程に関する知見は得られていない。 本研究では薄膜の成長について、初期過程における核形成機構の解明や単分子層の成長機構の解明を目的とし、まず基板上での単分子層が安定に観測できる系を探索した。その結果、表面にダングリングボンドを持たない層状物質基板上に分子性結晶であるC60、C70などのフラーレンを薄膜成長させた場合、これらの薄膜が単一配向のエピタキシャル膜を形成し、なおかつ基板上の単分子膜の形状を大気中での原子間力顕微鏡観察という比較的容易な手法で観測可能な系であることが分かった。この系を用いて様々な成長条件における薄膜の成長形態を観察することにより、エピタキシャル簿膜中の結晶軸配向、分子の基板上での拡散、核形成時の臨界核のサイズなど、薄膜成長に関してきわめて重要な幾つかの知見を得ることを試みた。 薄膜は分子線エピタキシー法によって作成し、基板および薄膜表面の結晶構造は反射高速電子線回折(RHEED)によってその場観察を行った。また、薄膜表面の形状は成長終了後に大気中で原子間力顕微鏡(AFM)により観察を行った。図1はMoS2基板上に成長したC60及びC70エピタキシャル膜の成長初期のRHEED像である。解析の結果、C60およびC70薄膜結晶は基板表面に沿って分子が稠密に配列した六回対称の格子を組んで成長することがわかった。薄膜中の分子間距離は基板結晶の格子定数とは無関係に成膜1層目からバルク結晶中の値と等しい値をとる。一方、基板と薄膜の結晶軸方向の関係は選択した基板と分子によって様々である。MoS2基板上におけるC60エピタキシャル膜は図2に示すように薄膜の軸配向を基板の軸方向に揃えて成長する。一方C70エピタキシャル膜の場合、薄膜の軸配向は基板の軸方向から30゜面内に回転した方向に揃う。解析の結果、MoS2基板上のC60、C70薄膜の配向は、それぞれ基板との格子不整合が小さくなる軸配向であることが分かった。いくつかの格子定数の異なる基板上での薄膜の配向観察を行った結果、基板表面での薄膜の配向は、基板・薄膜間の格子不整合を考えることによって説明できることがわかった。図3に基板と薄膜との格子定数の比afullerene/asubの値と薄膜の軸配向の関係を示す。図3で用いた基板上では、薄膜は比較的広いテラス上に二次元核生成によって成長を開始することがAFM観察より分かっており、このとき薄膜の軸配向は基板との格子不整合が最も小さくなる方向をとっている。 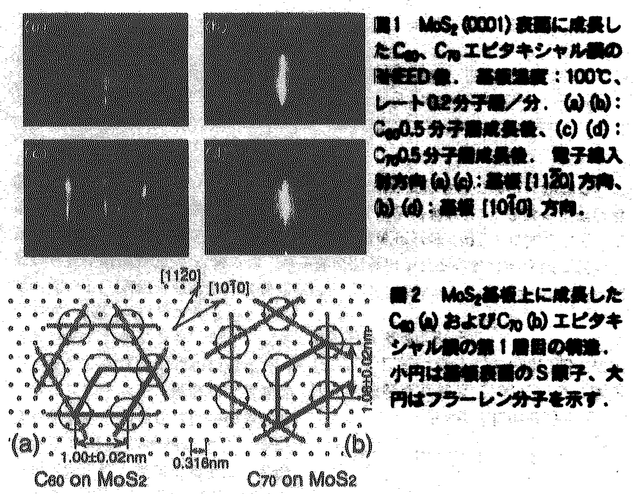 図表図1 MoS2(0001)表面に成長したC60、C70エピタキシャル膜のRHEED像.基板温度:100℃、レート0.2分子層/分.(a)(b):C60 0.5分子層成長後、(c)(d):C700.5分子層成長後.電子線入射方向(a)(c):基板[11 図表図1 MoS2(0001)表面に成長したC60、C70エピタキシャル膜のRHEED像.基板温度:100℃、レート0.2分子層/分.(a)(b):C60 0.5分子層成長後、(c)(d):C700.5分子層成長後.電子線入射方向(a)(c):基板[11 0]方向、(b)(d):基板[10 0]方向、(b)(d):基板[10 0]方向. / 図2 MoS2基板上に成長したC60(a)およびC70(b)エピタキシャル膜の第1層目の構造.小円は基板表面のS原子、大円はフラーレン分子を示す. 0]方向. / 図2 MoS2基板上に成長したC60(a)およびC70(b)エピタキシャル膜の第1層目の構造.小円は基板表面のS原子、大円はフラーレン分子を示す. 図3 C60エピタキシャル膜の格子定数と基板格子定数との比.()内は本実験より得られた基板結晶軸・薄膜結晶軸間の角度. 図3 C60エピタキシャル膜の格子定数と基板格子定数との比.()内は本実験より得られた基板結晶軸・薄膜結晶軸間の角度. AFMによる薄膜の表面形態観察では、選択する基板によってFrank-van der Merwe型(層状成長)からVolmer-Weber型(島状成長)まで、様々な成長形態が観測された。特にMoS2基板上のC60およびC70薄膜は層状成長の形態を示し、成膜1層目以降の各分子層の形態が明瞭に観察できるため、成長機構観察の系として適している。図4は一定の分子線強度(フラックス)における薄膜の成長初期の形態を各基板温度で測定したものである。AFM像中の最も暗い部分は基板表面である。その上に様々な形状の第1分子層と、三角形の形状をした第2分子層が成長している。第1分子層の形状に関しては、基板低温時には基板表面での核の数密度も高く1つ1つの核の形状は等方的であるが、基板高温時には核の密度が減少し、薄膜の結晶軸方向に枝を延ばした樹枝状構造を持つ。基板温度230℃においては側枝の成長も確認される。一般的に樹枝状成長は、過飽和状態における成長界面の濃度勾配が原因となって界面不安定が生じ、ある一定の結晶軸方向にそって急速に結晶が成長する、拡散律連結晶成長形態の一つとして考えられている。したがって今回得られたMoS2基板上のC60エピタキシャル膜第1層目の樹枝状構造は、C60分子の基板表面拡散が大きく影響していることが予想される。すなわち、MoS2基板上でのC60分子の拡散距離が長いために1つの核が取り込む分子数もそれだけ多くなることが、成長界面での過飽和の原因であると考えられる。この拡散距離の長さは、第1分子層の核密度が他の基板上の結果と比べて非常に小さいことからも同様に示唆される。一方、第2分子層以降の成長形態は安定面で囲まれた三角形の形状を持つ。これは第1分子層に比べて成長界面での過飽和度が低いことを示しており、核密度が第1分子層より高いことからも、拡散距離が短いことが予想される。第1分子層と第2分子層の成長形態の違いはMoS2基板表面のC60分子の拡散とC60分子層上のC60分子の拡散の違いが原因であると考えられる。  図4 種々の基板温度におけるMoS2基板上C60エピタキシャル膜AFM像.(フラックス:0.2分子層/分) 図4 種々の基板温度におけるMoS2基板上C60エピタキシャル膜AFM像.(フラックス:0.2分子層/分) 薄膜成長において基板上の成長核の表面密度は、薄膜の結晶性に影響を与える重要な情報であり、また様々な条件下での核密度の値を解析することによって、薄膜の成長機構やそれに必要ないくつかのパラメータを得ることができる。図5、図6にMoS2基板上のC60エピタキシャル膜に関して、飽和核密度の基板温度依存性と分子線強度依存性を示す。AFM観察から基板温度180℃の前後で第1分子層の形状が等方的なものから樹枝状へと変化することがわかっており、飽和核密度の温度依存性(図5)に関しても、この温度付近で第1分子層の傾向が変化していることは興味深い。金属や半導体薄膜の成長の場合にもこのような飽和核密度の基板温度依存性の変化は観測されており、この場合は臨界核のサイズの変化によって説明されている。臨界核のサイズは飽和核密度の分子線強度依存性から見積もることができる。図6から、基板温度低温時と高温時でグラフの傾きが異なっており、低温時には臨界核の大きさはC60分子4±1個程度、高温時にはC60分子数十個程度という結果を得た。一般的に臨界核の大きさがきいほど薄膜の成長形態は層状成長を示すことが分かっており、MoS2基板上のC60エピタキシャル膜における臨界核の大きさもAFM観察により確認された層状成長の成長様式を裏付けている。 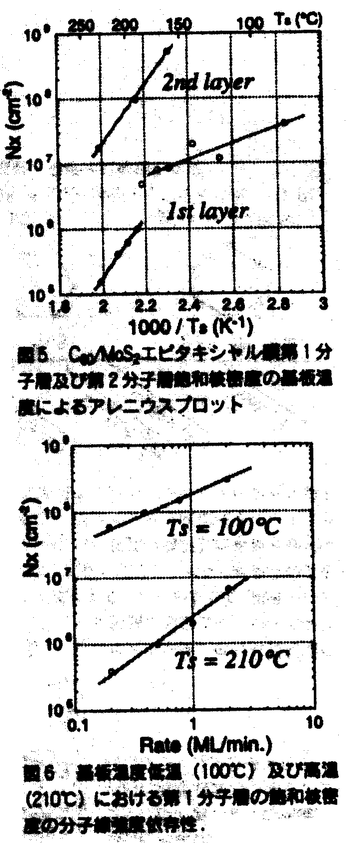 図表図5 C60/MoS2エピタキシャル膜第1分子層及び第2分子層飽和核密度の基板温度によるアレニウスプロット / 図6 基板温度低温(100℃)及び高温(210℃)における第1分子層の飽和核密度の分子線強度依存性. 図表図5 C60/MoS2エピタキシャル膜第1分子層及び第2分子層飽和核密度の基板温度によるアレニウスプロット / 図6 基板温度低温(100℃)及び高温(210℃)における第1分子層の飽和核密度の分子線強度依存性. 本研究では第2分子層目以降の多層膜の成長機構についても観察を行った。図7にMoS2基板上に成長したC60エピタキシャル膜の成長に伴う各分子層の成長の様子を観察したAFM像を示す。第1分子層は成長条件によって1つ1つの核が数十 mの大きさまで成長し得る。一方、多分子層膜の成長に関して第5分子層の成長まで追跡を行った結果、各分子層の成長様式は第2分子層以降全て等しく三角形の晶癖を持ち、各々の核のサイズも数m程度であることがわかった。このように基板に直接接した第1分子層のみが基板の影響を受けた成長様式を示し、第2分子層以降の基板に接しない層は一様に等しい成長様式を示す傾向は、各分子層の成長機構が直下の表面のみから影響を受けていることを示している。 mの大きさまで成長し得る。一方、多分子層膜の成長に関して第5分子層の成長まで追跡を行った結果、各分子層の成長様式は第2分子層以降全て等しく三角形の晶癖を持ち、各々の核のサイズも数m程度であることがわかった。このように基板に直接接した第1分子層のみが基板の影響を受けた成長様式を示し、第2分子層以降の基板に接しない層は一様に等しい成長様式を示す傾向は、各分子層の成長機構が直下の表面のみから影響を受けていることを示している。  図7 MoS2基板上C60エピタキシャル膜の膜厚による形態変化.(a):0.5分子層、(b):1.0分子層、(c):1.5分子層成長後.(基板温度:230℃観察領域:40m×40m) 図7 MoS2基板上C60エピタキシャル膜の膜厚による形態変化.(a):0.5分子層、(b):1.0分子層、(c):1.5分子層成長後.(基板温度:230℃観察領域:40m×40m) 以上、C60エピタキシャル膜において観測された各基板温度による臨界核サイズの変化や第1分子層の樹枝状成長など、成長初期過程に関する知見は金属や半導体の系で行われた薄膜成長観察では得られなかったものであり、本研究において初めて観測されたものである。MoS2基板上のC60エピタキシャル膜など、基板表面および薄膜がダングリングボンドを持たない系を選択することにより大気中での観測においても各分子層の成長機構が安定に観測が可能となり、新たな知見を得ることに成功した。 |