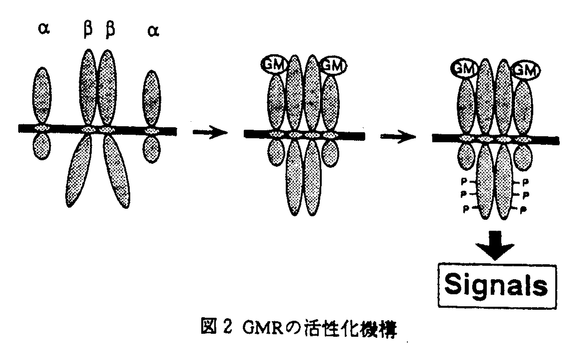
ヒトGM-CSFのレセプター(hGMR)は 鎖と 鎖と 鎖の二種のサブユニットによって構成され、高親和性かつ機能的なGMRを再構成するには両サブユニットが必須である。hGMRの活性化とそのシグナル伝達機構に関しては、鎖の細胞内領域の重要性についてのみ、欠失変異体を用いたhGMRの再構成実験によって詳細に解析されてきた。しかし、鎖の細胞内領域のシグナル伝達への関与、活性型レセブターの化学量論的な構造、およびサブユニット間の相互作用等に関しては不明であった。本論文では、これらの未解明の問題について、欠失変異体やキメラレセプターによるhGMRの再構成実験、および化学架橋剤を用いた生化学的な解析により検討を加えている。 鎖の二種のサブユニットによって構成され、高親和性かつ機能的なGMRを再構成するには両サブユニットが必須である。hGMRの活性化とそのシグナル伝達機構に関しては、鎖の細胞内領域の重要性についてのみ、欠失変異体を用いたhGMRの再構成実験によって詳細に解析されてきた。しかし、鎖の細胞内領域のシグナル伝達への関与、活性型レセブターの化学量論的な構造、およびサブユニット間の相互作用等に関しては不明であった。本論文では、これらの未解明の問題について、欠失変異体やキメラレセプターによるhGMRの再構成実験、および化学架橋剤を用いた生化学的な解析により検討を加えている。 本論文は三章からなる。第一章では、GM-CSFの作用、GMRの構造およびシグナル伝達について概説しており、本論文の序章として位置づけられている。 第二章は結果を示してあり、さらに三項に分かれている。第一項では欠失変異体を用いて鎖の細胞内領域のシグナル伝達への関与を検討し、以下の新たな知見を得ている。(1)鎖の細胞内領域の欠失により、細胞増殖、c-fosプロモーターの活性化、および鎖を含む細胞内蛋白質のチロシンリン酸化へのシグナルを伝達できなくなる。(2)鎖細胞内領域の中で、インターロイキン3(IL-3)、IL-5の鎖細胞内領域と高い相同性を有するN末端よりの29アミノ酸残基が、これらのシグナル伝達に重要である。(3)鎖と鎖はhGM-CSFに依存して複合体を形成し、この複合体形成に鎖細胞内領域の関与は認められない。以上の結果は、hGM-CSFのシグナル伝達に、鎖のみでなく鎖の細胞内領域も関与することを示しており、鎖の重要性を示す興味深い知見である。 第二項では、各サブユニットの細胞内領域の役割について検討するため、両サブユニット間で細胞内領域を交換したキメラサブユニット(/および/)を作製し、これらのキメラサブユニットおよび野生型のサブユニットを組み合わせた様々なキメラhGMRのシグナル伝達能について解析を行っている。その結果、鎖由来の細胞内領域のみを有するキメラhGMR(/,)が、hGM-CSFに依存した細胞増殖、初期応答遺伝子の発現、細胞内蛋白質のチロシンリン酸化を、野生型hGMRと同様のレベルで誘導できることを明らかにしている。 第三項では上記の結果からhGMR鎖が二量体を形成することを予測し、以下の事実を明らかにしている。(1)鎖が細胞外あるいは膜貫通領域を介して恒常的に二量体を形成している。(2)鎖二量体は、その単量体と同様、hGM-CSFに依存して鎖と複合体を形成する。(3)hGM-CSF依存的に鎖と相互作用した鎖二量体は、チロシン残基のリン酸化を受ける。 第三章では、本研究で得られた知見から各サブユニットの細胞内領域の役割、および活性型hGMRの構造について以下のように考察している。hGM-CSFのシグナルは、鎖由来の細胞内領域のみを有するキメラhGMR(/,)を通しても伝達される。このことは鎖がhGMRの主たるシグナル伝達分子であることを示している。しかし野生型のhGMRでは、鎖細胞内領域の欠失によりシグナルを伝達できなくなることから、通常は鎖は鎖との相互作用を通して活性化されること、キメラhGMRにおいては活性型の鎖の構造を模倣していることが示唆される。鎖は恒常的に二量体を形成しており、その活性化機構は他の多くのサイトカインレセプターに見られるリガンド依存的な二量体の形成では説明されえない。したがって、鎖細胞内領域との直接的相互作用による鎖の構造変化、あるいは鎖に結合する細胞内分子による鎖の修飾によって活性型の鎖に誘導されると推察している。さらにリガンドの構造およびそのレセプターとの結合様式を考え合わせ、活性型hGMRは、鎖二量体に二分子のhGM-CSFと二分子の鎖から構成されるというモデルを提唱している。 以上の研究は、hGMRの活性化機構に対して新たな知見を与えるものであり、さらにhGM-CSFのシグナル伝達の研究にも大きく貢献することが期待される。よって、論文提出者は、博士(理学)の学位を受けるのに十分な資格があるものと判断した。なお、本論文第二章は渡辺すみ子氏、宮島篤博士、新井賢一博士、横田崇博士との共同研究であるが、論文提出者が主体となって分析および検証を行ったもので、論文提出者の寄与が十分であると判断した。 |