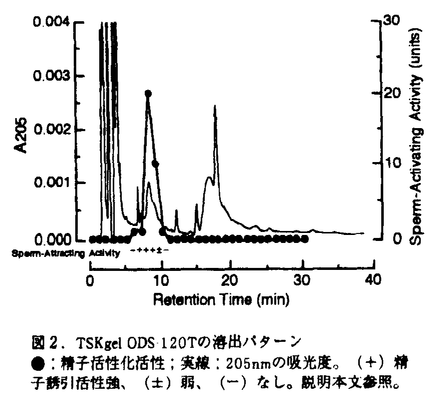
卵による精子の活性化及び誘引現象は、受精の確率を高めるシステムとして重要であり、また鞭毛運動の制御機構を研究していくうえでよい系である。本論文は原索動物であるホヤを用い、精子活性化・走化性機構の研究を行い3章からなっている。第1章は受精過程における精子活性化・走化性について、第2章は精子活性化・誘引物質の単離・精製、第3章は精子活性化・走化性の細胞情報伝達機構について述べられている。 第1章では、まずユウレイボヤ精子は海水中では運動性を持たないが、卵は受精時に精子を著しく活性化し、卵の植物極へ誘引すること、誘引物質は濾胞細胞等の付属細胞でなく卵本体から放出されることが明らかにされた。更に走化性を定量化する方法として極座標を用いた方法を開発し、受精時直後の卵形変化を境に卵の精子活性化・誘引活性能が失われることを明らかにした。又、cAMP分解酵素であるphosphodiestraseの阻害剤、theophyllineが精子活性化能を持つが、精子誘引能を持たないことを示し、この2つの活性は違う機構で制御されていると結論した。 第1章の結果から、ユウレイボヤ卵海水には強力な精子活性化、精子誘引活性がある。第2章では卵海水から精子活性化・誘引物質を精製した。ユウレイボヤ1500匹より得た1.2lの卵海水を凍結乾燥し、エタノール抽出及びクロロホルム-水系二層分配を行ない、粗精製標品を得、これを3種のHPLCカラム(SepPak C18、Shim-pak PRC-ODS、TSKgel ODS 120T)で順次精製し、205nmの紫外吸収に単一ピークを持つ活性標品を得た。この活性標品は精子活性化能と精子誘引能両方を備えていることから、2種の活性は同一物質によるものと結論され、この物質を精子活性化・誘引物質(Sperm-Activating and-Attracting Factor;SAAF)と命名した。このSAAFをガラス微小管の先端に充填し、精子懸濁液中に入れると、その周囲の精子は即座に活性化され、"chemotactic turn"と呼ばれるカーブを描きガラス管先端に集まる。 この標品の活性は煮沸30分、タンパク質分解酵素、グリコエンドペプチダーゼA、7種のレクチンの処理により影響を受けなかった。この結果はSAAFがタンパク質ではないこと、SAAF分子の中に糖鎖が含まれないかもしくは活性部位には関係していないことを示唆している。現在この物質の構造解析を試みている。 第3章では、精子活性化及び走化性の分子機構をSAAFを用い検討している。まずCa2+欠如海水中では精子はSAAFによって活性化、走化性を示さないが、Ca2+を加えると両方の活性が起こった。また、3種のCa2+チャネル阻害剤では、flunarizineが強い阻害作用を示した。このことはSAAFは細胞膜のT型Ca2+チャネルを開き、細胞外からCa2+が流入し、精子活性化・走化性を起こすことを示唆している。 一方、精子活性化能を持つtheophyllineはCa2+欠如海水中でも精子を活性化する。しかしこのtheophylline活性化精子でもCa2+欠如海水中ではSAAFに走化性を示さない。従ってcAMPは精子活性化には必要であるが精子走化性には必要ではない。また、精子内のcAMP量の変化をradioimmunoassay法で調べたところ、SAAFを加えただけではcAMP量は増大せず、Ca2+添加による運動活性化時にcAMP量が増大した。このことは流入したCa2+がアデニル酸シクラーゼを活性化し、cAMP量が増大してcAMPが精子活性化を起こすことを示している。 以上、ユウレイボヤ未受精卵では受精に至るまでの間に精子活性化と誘引の2つの現象を引き起こす物質、精子活性化・誘引物質(SAAF)が放出され精子に作用し、Ca2+とcAMPを必要とする精子活性化の経路と、Ca2+のみを必要とする精子走化性の経路を開くことが明かとなった。この研究は精子活性化・走化性の双方の細胞情報伝達機構を明らかにした最初の研究であり、今後SAAFの最終精製と構造決定、精子活性化におけるCa2+依存性アデニル酸シクラーゼやAキナーゼの関与、走化性におけるCa2+の作用機構の解明を通してさらなる研究の飛躍が期待される。 本論文の第1章は森沢正昭、稲葉一男博士、第2章は森沢正昭、稲葉一男、村田道雄博士、第3章は森沢正昭、石田克美、稲葉一男博士との共同研究であるが、論文提出者が主体となって分析及び検証を行なったもので、論文提出者の寄与が十分であると判断する。 |