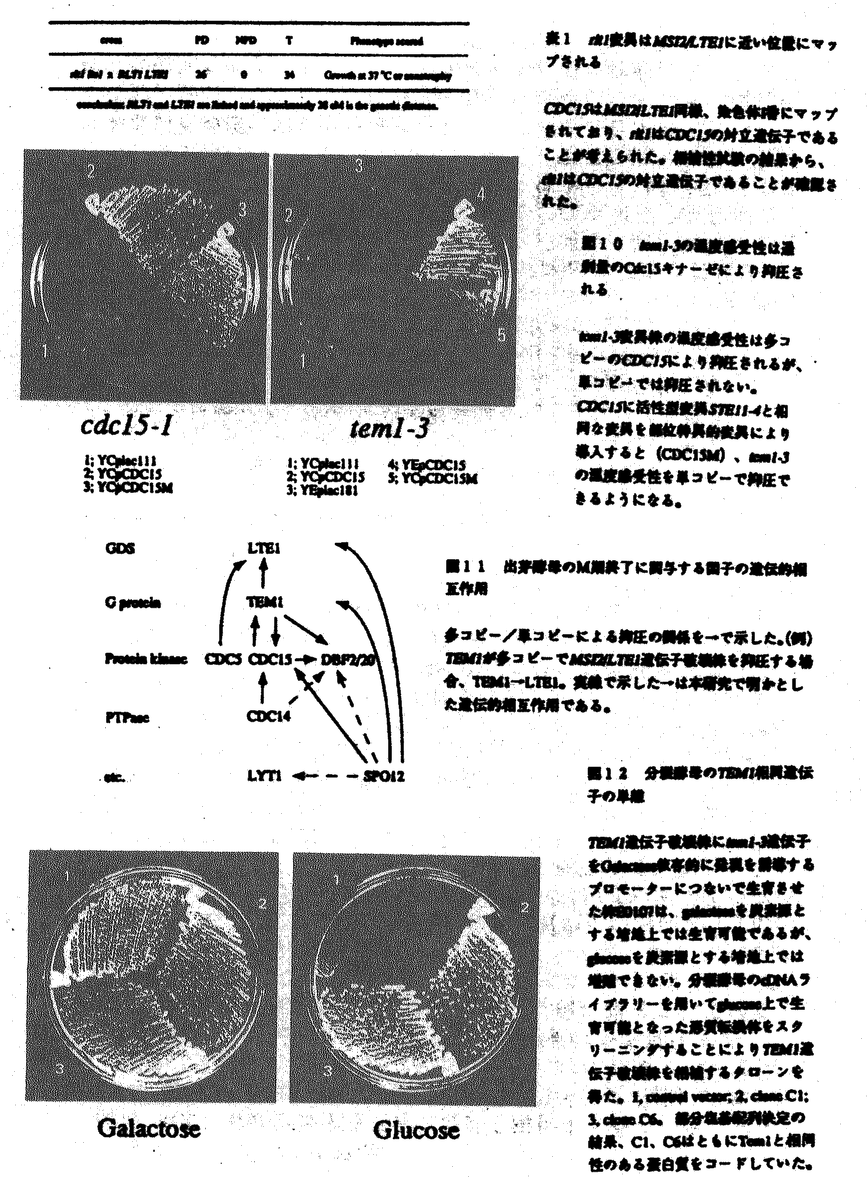
本論文は,ras-cAMP経路で働く遺伝子の研究中に見出されたMSI1遺伝子の破壊株が低温感受性を示し、制限温度下でM期を終了することが出来ないという現象の発見に端を発し、M期終了に係わる遺伝子群の発見とその遺伝的な相互作用を明らかにしたものである。本論文は4章からなり、第1章では出芽酵母のras-cAMP経路の遺伝子として分離したMSI2遺伝子の機能解析を行ない、第2章では低分子量GTPase遺伝子TEM1の発見とその機能を調べた。第3章では低温感受性msi2変異体の復帰変異の中で温度感受性になる変異体を分離し、それがcdc15変異であることを見出した。第4章ではM期終了に係わる遺伝子間の相互作用を検討し、それらの作用モデルを提案した。 真核生物が体細胞分裂及び減数分裂を開始するには、サイクリン依存性のセリン/スレオニンキナーゼ(CDK)の活性化が中心的な役割を果たす。活性化されたCDKはその標的蛋白質をリン酸化することにより、染色体凝縮、細胞骨格系の再構築、核膜の崩壊など、核分裂に必須の変化を引き起こすことが明かとなっている。CDKの活性の調節はM期の開始だけでなく、M期の維持及び終了のためにも重要と考えられる。染色体が分配されるまでCDKの活性は高い状態で維持され、G1期への移行には速やかにその活性が低下しなければならないからである。 細胞周期の研究は、酵母のcdc(cell division cycle)変異株の解析よりその制御機構が明かとなりつつあるが、M期終了の研究は他の生物種を含め、まだその制御因子及び制御機構について不明の点が多い。出芽酵母ではCDKが負に制御されないと細胞はM期を終了できずに増殖を停止する。従って、G2/M移行期同様、M/G1移行期においてもCDK活性の制御に関わる何らかの情報伝達系があることが予想される。私は、出芽酵母を用いた遺伝学的解析より、M期の終了を制御する低分子量G蛋白質を中心とした情報伝達系が存在することを明かにした。 1)MSI2/LTE1遺伝子の解析 酵母Ras蛋白質が制御する信号伝達系を解析するため、多コピーでira1変異株(Ras蛋白質のGTPase活性促進因子の欠損株)の熱ショック感受性を抑圧する因子MSI2を解析した。MSI2は多コピーで活性型RasをコードするRAS2val19の熱ショック感受性をも抑圧する。MSI2遺伝子の塩基配列を決定したところ、以前に報告されていたLTE1遺伝子と同一であった。しかし、報告されていた塩基配列には誤りがあり、正しくはMSI2/LTE1は1432アミノ酸をコードする遺伝子であった。Msi2はRas様低分子量G蛋白質の正の調節因子(GDS)であるCdc25,Scd25,Bud5と相同性がある。MSI2/LTE1遺伝子破壊株は低温感受性を示し、大きな芽を出したところで増殖を停止した。MSI2/LTE1遺伝子破壊株は、許容温度下でもチューブリンの脱重合剤であるベノミルに感受性を示した。MSI2/LTE1遺伝子破壊株が制限温度下で細胞周期のどの時点で増殖を停止しているか、さらに詳しく調べるため、細胞を固定し、DAPIによりDNAを、間接蛍光抗体法によりチューブリンを染色した。その結果、核の複製及び娘細胞への移行は終了していたが、両核を結ぶスピンドルは残ったままであった。従って、Msi2/Lte1は細胞周期の核分裂終期で機能していることが分った。Msi2はRas様低分子量G蛋白質の正の調節因子と考えられることから、M期終了時において低分子量G蛋白質による情報伝達経路が存在すると考えられた。そこで、M期終了の制御機構の解明を目的に、MSI2/LTE1遺伝子破壊株の多コピー抑圧遺伝子及び復帰突然変異株を取得し、解析を行った。 2)低分子量G蛋白質をコードするTEM1遺伝子の解析 MSI2/LTE1遺伝子破壊株の低温感受性の多コピー抑圧遺伝子としてTEM1を単離した。相同性検索の結果から、TEM1は新規のファミリーに属する低分子量G蛋白質をコードすることが明かとなり、実際にGST-Tem1融合蛋白質は特異的にGTP Sと結合することが分った。TEM1遺伝子は増殖に必須で、その温度感受性変異株tem1-3は制限温度下で核分裂終期で増殖を停止した。これはMSI2/LTE1遺伝子破壊株と同じ表現型であり、Msi2/Lte1がTem1の正の調節因子の一つである可能性を示唆する。通常、細胞周期のM期に開始に必要なCDK活性は、核分裂後期で不活化される。CDKが核分裂後期で不活化されないと細胞は核分裂終期で増殖を停止し、M期を脱出できない。一つの可能性として、tem1-3は制限温度下でCDKを負に制御できないため核分裂終期で増殖を停止していることが考えられた。そこで、tem1-3の増殖停止時のCDK活性を測定したところ、細胞が核分裂後期を終了しているにもかかわらず、高い状態にあった。また、tem1-3は許容温度下でも、過剰のCLB2(B型サイクリンをコードする)を発現させることでCDKの活性を上昇させると核分裂終期で増殖を停止した。従って、Tem1はCDK不活化することでM期の終了を制御していることが考えられた。 Sと結合することが分った。TEM1遺伝子は増殖に必須で、その温度感受性変異株tem1-3は制限温度下で核分裂終期で増殖を停止した。これはMSI2/LTE1遺伝子破壊株と同じ表現型であり、Msi2/Lte1がTem1の正の調節因子の一つである可能性を示唆する。通常、細胞周期のM期に開始に必要なCDK活性は、核分裂後期で不活化される。CDKが核分裂後期で不活化されないと細胞は核分裂終期で増殖を停止し、M期を脱出できない。一つの可能性として、tem1-3は制限温度下でCDKを負に制御できないため核分裂終期で増殖を停止していることが考えられた。そこで、tem1-3の増殖停止時のCDK活性を測定したところ、細胞が核分裂後期を終了しているにもかかわらず、高い状態にあった。また、tem1-3は許容温度下でも、過剰のCLB2(B型サイクリンをコードする)を発現させることでCDKの活性を上昇させると核分裂終期で増殖を停止した。従って、Tem1はCDK不活化することでM期の終了を制御していることが考えられた。 3)MSI2/LTE1の温度感受性復帰突然変異株rlt1の解析 msi2/lte1遺伝子破壊株の低温感受性の復帰突然変異株rlt1を単離した。得られた変異株msi2 rlt1は低温で増殖可能であるが、逆に高温では増殖できなくなった。rlt1変異株は核分裂終期で増殖を停止した。これは、msi2/lte1遺伝子破壊株、tem1-3変異株と同じ表現型である。また、rlt1の5つの対立遺伝子(rlt1-1からrlt1-5)は、温度感受性において劣性であり、lte1の低温感受性の抑圧は優性であった。従って、RLT1遺伝子産物は低温下でgain-of-function、高温下でloss-of-functionであると考えられる。遺伝学的解析からRLT1はMAPKKK類似のプロテインキナーゼをコードするCDC15の対立遺伝子であることが分かった。そこで、TEM1とCDC15との遺伝的相互作用を調べた。温度感受性変異株tem1-3は多コピーのCDC15により抑圧される。CDC15は多コピーでTEM1遺伝子破壊株の致死性をも抑圧することから、Tem1とCdc15の機能が互いに重複しているか、共通の情報伝達系でCdc15がTem1の下流で働いているか、が考えられる。 4)M期終了を制御する因子の遺伝的相互作用 CDC15の下流の因子を得る目的で、rlt1-1の温度感受性変異株の単コピー抑圧遺伝子及び多コピー抑圧遺伝子を単離した。単コピー抑圧遺伝子として得られた遺伝子はチロシンホスファターゼをコードするCDC14であった。部位特異的変異を導入することにより、チロシンホスファターゼとして活性を持たないと考えられるCDC14の対立遺伝子を作製した。これらの対立遺伝子はcdc14-1の温度感受性を抑圧できないが、rlt1-1の温度感受性は抑圧することができた。このことから、単コピーのCDC14によるrlt1-1変異の抑圧にチロシンホスファターゼの活性が関与せず、CDC14遺伝子産物がチロシンホスファターゼ以外の機能を持つ蛋白質であることが示唆された。多コピー抑圧遺伝子として得られた遺伝子はSPO12と同一であった。SPO12は減数分裂に必須の遺伝子で、その遺伝子破壊株は減数分裂を行う際、減数第一分裂をスキップして減数第二分裂を開始することが報告されている。SPO12はtem1-3の多コピー抑圧遺伝子としても分離されており、MSI2/LTE1遺伝子破壊株をも抑圧することから、体細胞分裂のM期終了時においてもなんらかの機能を有していることが考えられる。 以上のように本論文は、これまで未知であったM期終了の分子機構の解明に道筋を付けるものである。ここで提出されたモデルを基礎に、さらに生化学的研究、細胞生物学的研究を進めてゆくことのより細胞分裂の終了の全体像を理解できるようになると思われる。本論文は共著であるが、実験計画の立案と執行は申請者自身によるもので、他のものは実験指導者である。以上の評価に基づき、本研究は博士(理学)の学位に十分値するものであることが、審査委員全員一致により認められた。 |