| 審査要旨 | | 本論文は,昆虫フェロモンに関する研究で,第一章,第二章の第一部,第三章,第四章の第二部により構成されている。第一部は,フェロモンの化学構造と生物活性の相関を調査するために行った数種の昆虫フェロモンの化学合成研究についてであり,第二部ではフェロモンの生合成にあずかる神経ペプチドPBANの遺伝的支配を解明するために行ったPBAN遺伝子の連関分析について述べている。 多くの種の昆虫は,フェロモンと呼ばれる化学物質を用い,同種の個体間で相互に密接な情報交換を行っていることが知られているが,まず第一章で,フェロモンの定義・分類,これまでの昆虫フェロモン関する研究の歴史を概説した後,第二章では3種の昆虫フェロモンの化学合成について述べている。 まず,アリの一種(Crematogaster auberti)の働きアリの道しるベフェロモンとして単離された2-ドデカノールの両鏡像体を図示の通り合成した。 まず光学活性な3-ヒドロキシブタン酸エチル(R)-1より出発し,水酸基をTHPエーテルとして保護し,還元,トシル化した。得られたトシレート2は,Grignardカッブリングにより3へと導き,3は,脱保護して(R)-2-ドデカノールとした。光学純度は100%であった。 (S)-体も同様に(S)-1より合成し,合成した両鏡像体の生物試験の結果,(R)-体が(S)-体の約1000倍の活性を有することが明らかとなった。 次にアリの一種Lasius nigerのフェロモン構成成分であるが,立体構造不明の3種のラクトン,A,B,Cを共通の出発原料である(R)-シトロネロール5より合成した。7→8の熱力学的支配下でのヨードラクトン化を鍵反応としている。これにより,高光学純度のA〜Cの合成が達成され,生物学者への研究用サンプルの供給が可能となった。 もう1つの合成は,蛾の一種(Nepticula malella)の性フェロモン,(S,Z)-6,8-ノナジエン-2-オールについてである。このものも,(S)-1より図示の様な炭素鎖延長を経て合成された。 第二部では,性フェロモン生合成の遺伝学的解析を行っている。 まず第三章でカイコの性フェロモンであるポンビコールの生合成を活性化するPBANに関してこれまでになされてきた研究について概説した後,第四章においてフェロモン生合成の遺伝的背景を解明するために,カイコのPBANをコードする遺伝子の品種間多型を探素し,染色体上の座位を決定した。また,カイコには多くの休眠性の遺伝的変異が存在するが,カイコのPBAN遺伝子はPBANと同時に胚休眠を誘導する休眠ホルモン(DH)をコードしているため,これらの休眠性変異とDH-PBAN遺伝子の関連性も調べた。 カイコ20品種からDNAを調製し,PCR法を利用してイントロンのサイズ多型を探索した。その結果,DH-PBAN遺伝子(Dh)には少なくとも3つの複対立遺伝子があることが判明し,それらをDhA1,DhA2並びにDhBと名付けた。さらに,DhA1,DhA2,DhB遺伝子を持つ系統の交雑,正逆交雑,戻し交雑試験により,Dh遺伝子が常染色体の上に座位するものと推定した。また,EKp(第6連関群),pe-re(第5連関群),ch(第13連関群),pM(第2連関群),Ze(第3連関群),L(第4連関群)の6種のマーカー遺伝子と独立であることもわかった。 また,w30系統の持つ黒蛹bp(第11連関群)と連関関係を示す分離が得られたことからDhがbp遺伝子と連関関係にあり,第11連関群に所属すると推定し,その確認としてbp以外の第11連関群の突然変異遺伝子である瘤蚕K,火傷蚕Buとの連関関係を検定交雑によって調査した。その結果からもDh遺伝子がKとBuと連関し,11連関群へ所属していることがわかった。 次にDH-PBAN遺伝子の座位を決定するため,KとBuを基準として3点交雑実験を行い,Kが23.2に,Buが28.7に,mpが51.8に,それぞれ座位することから,Dh-K-Bu-mpの順序に各遺伝子が配列すると推定し,Dhは第11連関群の-2.2cMに占座すると結論した。 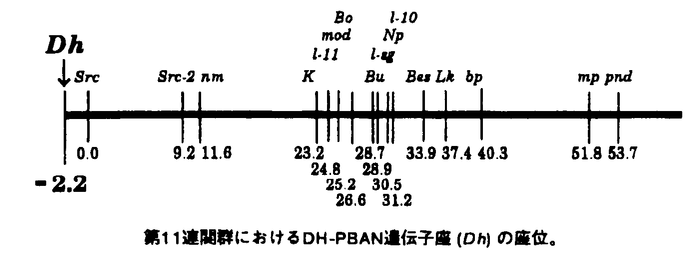 第11連関群におけるDH-PBAN遺伝子座(Dh)の座位。 第11連関群におけるDH-PBAN遺伝子座(Dh)の座位。 以上本論文は,立体制御された化学合成により昆虫フェロモンを合成し,また,性フェロモンの生合成を制御する遺伝子について,その構造的変異と座位を明らかにしたものであり,学術上貢献するところが少なくない。よって審査員一同は,申請者に博士(農学)の学位を授与してしかるべきと判定した。 |



