学位論文要旨
| No | 111289 | |
| 著者(漢字) | 西田,洋巳 | |
| 著者(英字) | ||
| 著者(カナ) | ニシダ,ヒロミ | |
| 標題(和) | 高等菌類の系統進化に関する研究 | |
| 標題(洋) | ||
| 報告番号 | 111289 | |
| 報告番号 | 甲11289 | |
| 学位授与日 | 1995.03.29 | |
| 学位種別 | 課程博士 | |
| 学位種類 | 博士(農学) | |
| 学位記番号 | 博農第1580号 | |
| 研究科 | 農学生命科学研究科 | |
| 専攻 | 応用生命工学専攻 | |
| 論文審査委員 | ||
| 内容要旨 | 菌類の系統進化の解明は主として形態学の研究から進められてきた。しかし、多様化したなかにも、単純で、しばしば収斂する形態、乏しい化石資料は本研究の進展を阻害している。遺伝子やタンパク質の比較により系統進化を探る分子系統学は菌類の系統進化の研究を未来あるものとした。さらに、分子生物学の発展にともない分子進化学が菌類における形態の差異を遺伝子レベルで解明しようとしている。本研究では形態学では明確にならなかった高等菌類に対し分子系統学的解析を行い、形態学、生理生化学による解析と比較した。また、グループIイントロンの分布機構を解明する目的で、分子進化学的解析を行った。 高等菌類の系統は有性時代において外生の担子胞子をつくる担子菌類と内生の子嚢胞子をつくる子嚢菌類の系統群に分けられる。どのようにして高等菌類は子嚢菌類と担子菌類に分岐し、さらにそれぞれの系統群において、単細胞の酵母から菌糸をはりめぐらした多細胞のものにまで多様化したのであろうか。本研究の目的の1つはこの問題を解くことであった。 種々の高等菌類は、核小サブユニットリボソームRNA(18S rRNA)に基づき系統樹を作成すると、明確に担子菌類と子嚢菌類の系統群に分かれた。有性胞子形成様式と18S rRNAは互いに独立したものである。しかし、高等菌類を2つのグループに分ける点で完全に一致した。18S rRNAが高等菌類の系統を反映しているからである。18S rRNA比較に基づいた子嚢菌類系統群の中に子嚢菌類の共通祖先から最初に分岐してきた系統群を見出し、古生子嚢菌類(Archiascomycetes)として提唱した(1,2,図1)。子嚢菌類における他の2つの系統群、真正子嚢菌類(Euascomycetes)と半子嚢菌類(Hemiascomycetes)はそれぞれ共通の特徴的な形態を持つ。真正子嚢菌類は、生活環の大部分を菌糸状で過ごし、半子嚢菌類は酵母で過ごす。これに対し、古生子嚢菌類は形態的に共通のものが見出せず多系統であることを示唆している。 ゼンマイ腫葉病菌Mixia osmundaeはその不十分な形態観察にもとづき子嚢菌類に分類され、菌類学者の間で80年間以上にもわたって子嚢菌類の1つであると信じられてきたが、18S rRNA塩基配列比較によると担子菌類系統に属した。梅雨期の直前、ゼンマイが葉を広げる頃に静岡山中にてゼンマイ腫葉病を見つけ、そこからM.osmundaeを分離した。改めて、その18S rDNAの塩基配列の決定および微細構造を観察したところ典型的な担子菌類の特徴を示した。そこで、M.osmundaeを子嚢菌類から担子菌類へ移した(3)。これは日頃分子系統には無関心な菌類学者に強烈なインパクトを与えた。 植物寄生菌類Protomycesの18S rDNA領域にグループIイントロンの存在を示した(4,5)。グループIイントロンは自己スプライシングRNAの1つである。また、一部のグループIイントロンは移動性をもつ。その分布は生物の系統を反映せず、異種生物間における水平移動が考えられた。オニタビラコ浮腫病菌Protomyces inouyeiに存在した2つのグループIイントロンの1つはジシバリ浮腫病菌P.lactucae-debilisに存在したグループIイントロンと挿入位置が一致した上、塩基配列において98.5%の相同性を示した。本イントロンの祖先はP.inouyeiとP.lactucae-debilisの共通祖先にすでに存在していたことを示す。 グループIイントロンはP,Q,R,Sの4つの保存領域を持っている。PとSの塩基配列に基づきプライマーを作成し、PCRを行った結果、オニタビラコYoungia japonicaにおいて増幅産物を得た。他方、ジシバリlxeris debilisでは増幅産物は得られなかった。オニタビラコの増幅産物の塩基配列を調べたところ、4つの異なるグループIイントロンの塩基配列を得た。その中の1つはProtomyces inouyeiに特異的なグループIイントロンの塩基配列と一致した。さらにその挿入位置はProtomyces inouyeiのそれとは異なることが明らかとなった。 すなわち、Protomyces inouyeiに存在した2つのグループIイントロンの分布機構はそれぞれ異なっている。1つはProtomyces inouyeiとP.lactucae-debilisが進化上分岐する以前の共通祖先にすでに存在し、もう1つは分岐の後、P.inouyeiの宿主植物オニタビラコから水平移動し、固定されたことを示す(6)。また、グループIイントロンは藻類に多く発見されている。植物の色素体、その祖先と関係が深いシアノバクテリアでも見つかっている。よって、現在後生動物以外の多くの生物で見つかっているグループIイントロンと藻類のグループIイントロンは深いかかわりがあると考えられる。グループIイントロンはウイルスにも発見されていることからその分布機構はかなり複雑であると考えられる。以上のことから、図2のような過程を経てProtomycesにグループIイントロンが存在するようになったと考えられる。 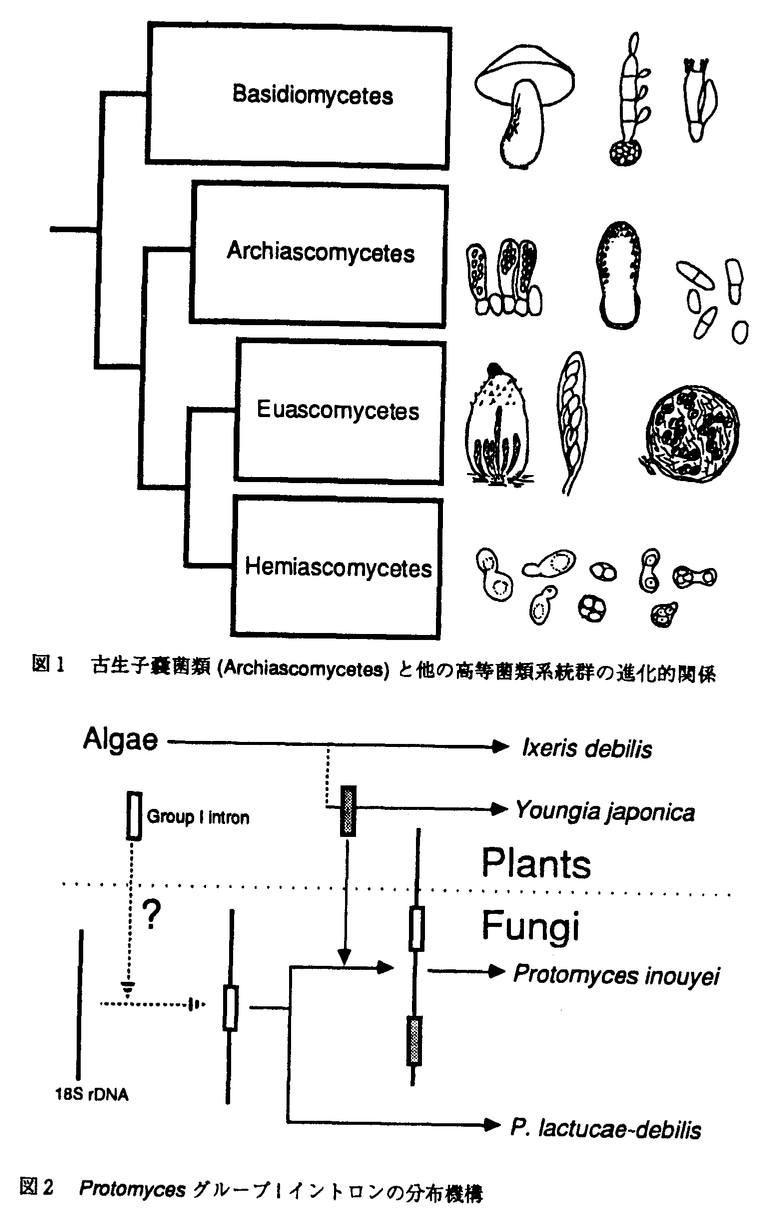 約4億年前に高等菌類の祖先は担子菌類と子嚢菌類の系統にわかれた。その後、それぞれの系統群において多様化し、現生高等菌類になった。無性時代において出芽増殖する酵母は担子菌類にも子嚢菌類にも見られるが、それらの祖先は約4億年前まで遡らない限り存在しない。形態的収斂の結果である。しかし、現菌類分類体系は形態学に基づいている。18S rRNAの塩基配列に基づく高等菌類の系統は、その祖先から現生高等菌類に至るまでの道筋を示している。形態的特徴はこの系統樹には反映されないにもかかわらず、主要系統群とその形態的特徴はよく一致した。形態上の進化の過程が追跡できる。今後、形態形成のための情報がいかに遺伝子に刻まれているか明らかとなれば高等菌類の系統学は完成するだろう。 単独で生きている生物は存在しない。寄生や共生はその顕著な例である。本研究ではアミノ酸やタンパク質だけでなく、異種生物間を遺伝子が水平移動し、固定される現象を示した。一方的な依存関係である寄生から相互依存関係である共生へ変化するきっかけが、遺伝子の水平移動にある可能性が高い。 | |
| 審査要旨 | 菌類の系統進化の解明は,主として形態レベルの研究から進められてきた。しかし,多様化した中にも,単純で,しばしば収斂する形態,乏しい化石資料はこの研究分野の進歩を阻害している。遺伝子やタンパク質の比較により系統進化を探る分子系統学の登場は,菌類の系統進化の研究を客観性のある科学に高めた。さらに,分子生物学の発展にともない分子進化学は菌類における形態の差異を遺伝子レベルで解明しようとしている。本論文は,従来の比較形態学的手法では推定の域を出なかった高等菌類の系統進化について分子レベルから解析を行い,主要な表現形質と比較解析してその大筋を明らかにし,さらに18SリボンームDNA中に発見したGroup I Intronについて分子進化的解析を行い,その分布機構について考察したものである。 第1章(序論)で,研究の背景と意義を概説した後,第2章で,Taphrina,Protomyces,Saitoella属の代表的な5種の核18S rDNAの塩基配列を決定し,既知データを加えて系統樹を作成し,多面的に解析した結果について述べている。高等菌類は子嚢・内生子嚢胞子を形成する子嚢菌類と担子器・外生担子胞子を形成する担子菌類の2つの大きな系統群に分けられる。さらに,子嚢菌類は3つの主要系統群に分かれ,その共通祖先から最初に分岐してきた主要系統群はTaphrina,Protomyces,不完全酵母Saitoella,分裂酵母Schizosaccharomyces,カリニ肺炎菌Pneumocystisを含み,古生子嚢菌類(Archiascomycetes)と命名,提唱した。子嚢菌類における他の2つの主要系統群は半子嚢菌類と真正子嚢菌類はそれぞれ単系統であることを明確に示した。しかし,古生子嚢菌類は形態レベルで共通の分類形質が見い出せず多系統であることを示唆した。 第3章では,ゼンマイ腫葉病菌Mixia osmundaeの系統分類学的位置について述べている。本菌は,その形態的特徴から原記載以来80年以上にもわたって子嚢菌類の一種であると菌学者の間で信じられてきた。本種の保存株と新たに野外で採取したゼンマイ腫葉病菌から分離した株について,18S rDNA塩基配列を決定,系統解析したところ,定説に反して担子菌類系統に帰属した。そこで,新鮮な試料について微細形態学的観察を行い,胞子は,従来子嚢と考えられていた胞子形成細胞から外生的,分芽的,同調的に生ずることを見いだした。従って,得られた分子と形態両形質のデータは完全に一致し,M.osmundaeは子嚢菌類から担子菌類に移籍された。 第4章では,植物寄生菌類Protomyces属2種の18S rDNA領域にGroup I Intronを見いだし,それらの全塩基配列を決定し,その進化学的意義について述べている。すなわち,オニタビラコ浮腫病菌P.inouyeiに存在した2つのGroup I Intronの一つはジシバリ浮腫病菌P.lactucaedebilisに存在したGroup I Intronと挿入位置が一致したうえ,塩基配列において98.5%の相同性を示した。このことは,本Intron の祖先はP.inouyeiとP.lactucae-debillsの共通祖先にすでに存在していたことを示すものである。さらに,宿主植物についてもPCR法による検出を試みた結果,オニタビラコYoungia japonicaにおいて増幅産物を得た。その塩基配列を調べたところ,4つの異なるGroup I Intronの塩基配列を見いだした。その中の一つはP.inouyeiに特異的なGroup I Intronの塩基配列と一致した。以上のデータを解析して,P.inouyeiに存在した2つのGroup I Intronの分布機構はそれぞれ異なっていることを示した。 第5章は総合考察である。現菌類分類体系の骨格は形態形質に基づいている。分子形質としての18S rRNA遺伝子の塩基配列に基づく高等菌類の系統樹は,その祖先から現生高等菌類に至る進化の道筋を示している。形態形質はこの系統樹には,反映されないにもかかわらず,主要系統群とその主要な形態形質はよく一致した。また,本研究でProtomyces属菌で発見した18S rRNA遺伝子領域のGroup I Intronの異種生物の間での移動について考察を加え,P.inouyeiに存在したGroup I Intronの分布機構はそれぞれ異なっていると結論した。すなわち,1つはP.inouyeiとP.lactucae-debilisが進化上分岐する以前に共通祖先にすでに存在し,他の1つは分岐後,P.inouyeiの宿主植物オニタビラコから水平移動し,固定されたとの説を導いた。 以上本論文は,これまで推定の域を出なかった高等菌類の系統進化の問題に分子レベルから光を当て,その大筋を明らかにし,さらに異種生物間における遺伝子の水平移動の可能性を提示したもので,学術上貢献するところが少なくない。よって審査員一同は,本論文が博士(農学)の学位論文として価値あるものと認めた。 | |
| UTokyo Repositoryリンク | http://hdl.handle.net/2261/54470 |