| 審査要旨 | | イネ根圏に共生する窒素固定細菌を応用して化学窒素肥料と置き換えようという試み,即ち「生物窒素肥料」の開発は,経済的ならびに環境的な観点より,現在の農業における極めて重要な課題であゐ。 本論文はイネ根圏より分離された窒素固定細菌Klebsiella oxytocaならびにAzospirillum lipoferumに注目し,その窒素固定能の増強を目的として窒素固定遺伝子の発現制御機構について研究した結果を述べたものである。 第1章序論に続いて,第2章では,K.oxytocaの改良菌株をイネ根圏に接種してその生育に及ぼす影響を検討した。その結果,単一の菌のみを多量に接種しても,急速に死滅してしまうことが判明した。したがって,根圏における菌の比率をなるべく変えないよう,複数の窒素固定菌を改良し,同時に用いることが理想的な窒素固定系の確立のために必要であると考えられた。そこで以降A.lipoferumの改良の第一段階として同菌におけるNifA蛋白を介した窒素固定遺伝子の発現制御機構の解明を行うこととした。 第3章では,A.lipoferumのnifA遺伝子の単離と塩基配列の決定について述べている。A.lipoferum FSのゲノムDNA上よりnifA遺伝子を約2.8kbのSalI断片としてクローン化した。塩基配列を決定したところ,624アミノ酸をコードするORFが見出され,同ORFの推定アミノ酸配列はK.oxytocaのNifA蛋白に対して約40%の一致を示した。他の窒素固定菌のNifA蛋白と一次構造を比較したところ,蛋白の中央部に位置する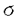 54-interaction domainはA.lipoferumにおいてもアミノ酸配列上250番から450番の領域によく保存されていた。また,ヘリックスターンヘリックスタイプのDNA binding motifをふくむDNA binding domainもC末端部によく保存されていた。 54-interaction domainはA.lipoferumにおいてもアミノ酸配列上250番から450番の領域によく保存されていた。また,ヘリックスターンヘリックスタイプのDNA binding motifをふくむDNA binding domainもC末端部によく保存されていた。 第4章ではNifA蛋白の標的であるnifプロモーターの取得を目的として,A.lipoferumのnifH遺伝子の単離と塩基配列の決定を行った。A.lipoferum FSのゲノムより約2.7kbのSalI-HincII断片としてnifH遺伝子をクローン化し,塩基配列を決定した。その結果,293アミノ酸をコードするORFが見出され,コードするアミノ酸配列はA.brasilenseのNifH蛋白のものと完全に一致した。また,この断片にはnifD遺伝子の一部も含まれており,nifHとオペロンを構成すると考えられた。 第5章ではA.lipoferumのnifAとnifH遺伝子の発現をノザンハイブリダイゼーション実験により解析した。アンモニアを含む培地で好気的に培養したA.lipoferumを無窒素培地に移し,微好気的条件で培養を続け,経時的にサンプリングを行いRNAを調製した。このRNAに対し,nifAの中央部約1kbのPstI断片,nifH中央部より下流全領域を含む約1kbのXhoI-HindIII断片をプローブとしたハイブリダイゼーションを行った。その結果,A.lipoferumにおいてnifA遺伝子の転写はアンモニアの存在下では抑制されており,無窒素培地において培養開始後4時間の間誘導を受けることを明らかにした。一方,nifHプローブを用いた実験により,nifH遺伝子の転写は無窒素培地でしかも微好気的条件下でのみ誘導を受け,無窒素培地での培養開始後4から6時間後に転写レベルが最も高くなることを明らかにした。 第6章ではNifA蛋白の機能を,A.lipoferumのnifA遺伝子を大腸菌において発現させ,同時に導入したnifH遺伝子の転写の活性化が起こるか否かを指標に解析した。lacプロモーターの下流にnifA遺伝子を連結したプラスミドとnifHプロモーターの下流にlacZ遺伝子を連結したプラスミドを同時にlac-の大腸菌に導入したところ,好気的な条件では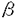 -ガラクトシダーゼ活性は発現せず,低酸素分圧あるいは嫌気的条件でのみ活性が発現した。即ち,低酸素分圧下においてのみNifA蛋白がnifH遺伝子の転写を活性化することを明らかにした。また,K.oxytocaのnifA遺伝子を用いて同様の実験を行ったところ,酸素分圧にかかわらずnifH遺伝子の転写の活性化が観測された。すなわち,K.oxytocaではNifL蛋白が酸素分圧のセンサーとなってNifA蛋白による窒素固定遺伝子の発現を制御するのに対し,A.lipoferumにおいてはNifA蛋白自身がセンサーとなって酸素分圧に応じた窒素固定遺伝子群の制御を行なっている可能性をはじめて示した。 -ガラクトシダーゼ活性は発現せず,低酸素分圧あるいは嫌気的条件でのみ活性が発現した。即ち,低酸素分圧下においてのみNifA蛋白がnifH遺伝子の転写を活性化することを明らかにした。また,K.oxytocaのnifA遺伝子を用いて同様の実験を行ったところ,酸素分圧にかかわらずnifH遺伝子の転写の活性化が観測された。すなわち,K.oxytocaではNifL蛋白が酸素分圧のセンサーとなってNifA蛋白による窒素固定遺伝子の発現を制御するのに対し,A.lipoferumにおいてはNifA蛋白自身がセンサーとなって酸素分圧に応じた窒素固定遺伝子群の制御を行なっている可能性をはじめて示した。 以上本論文は,A.lipoferumにおけるNifA蛋白による窒素固定遺伝子の発現制御機構について研究し,その結果同菌のNifA蛋白はK.oxytocaには見られない独自の機能を持つことを明らかにしたものであり,応用的研究への展望を示すとともに,学術上貢献するところが少なくない。よって審査員一同は,本論文が博士(農学)の学位論文として価値あるものと認めた。 |