| 審査要旨 | | 本論文は,好熱性放線菌Thermoactinomyces vulgarisの -アミラーゼの基質特異性の解析と応用について述べたもので5章より成る。アミラーゼは,食品工業で最も需要が高い酵素であり,タンパク質工学的手法を用いて,より有用な酵素に改変するという研究は,大きな成果が期待されている。本研究の材料である,好熱性放線菌T.vulgaris R-47の生産する-アミラーゼ(TVA)は,通常の-アミラーゼが分解することのできない,多糖プルランを分解するという特徴的な基質特異性を持つ酵素である。T.vulgarisは,このような性質を持ちながら,アミノ酸配列が異なる二種類のアミラーゼ,TVAIとTVAII,を生産している。本研究ではこのような特徴的な酵素の比較により,アミラーゼの分子認識機構を解析することを目的として,以下の研究を行っている。 -アミラーゼの基質特異性の解析と応用について述べたもので5章より成る。アミラーゼは,食品工業で最も需要が高い酵素であり,タンパク質工学的手法を用いて,より有用な酵素に改変するという研究は,大きな成果が期待されている。本研究の材料である,好熱性放線菌T.vulgaris R-47の生産する-アミラーゼ(TVA)は,通常の-アミラーゼが分解することのできない,多糖プルランを分解するという特徴的な基質特異性を持つ酵素である。T.vulgarisは,このような性質を持ちながら,アミノ酸配列が異なる二種類のアミラーゼ,TVAIとTVAII,を生産している。本研究ではこのような特徴的な酵素の比較により,アミラーゼの分子認識機構を解析することを目的として,以下の研究を行っている。 序章で本研究の意義について概説した後,第1章では,TVAIIの大量発現系の構築,および結晶化について述べている。本研究では,TVAII遺伝子由来のSD配列を利用して,TVAIIの発現ベクターを構築することにより,培地1lあたり100mg以上のTVAIIを得る系を確立することができた。また,このようにして大量に得られたTVAIIの結晶化に成功し,X線構造解析に耐えうるものであることを明らかにした。 第2章では糖転移反応の応用と解析について述べている。本研究では,TVAの糖転移反応を利用したオリゴ糖の酵素合成とその解析を行った。プルランとグルコースを出発原料とすると,加水分解産物であるパノースのほかに,パノースにグルコースが(1→4)結合で転移した四糖42-isomaltosylmaltose(IMM)と(1→6)結合で転移した四糖42-isomaltosylisomaltose(IMIM)が生成する。本研究ではIMIMの分離精製に成功した。また,この合成においてTVAIIはこれらの糖転移生成物を大量に生成するが,TVAIは少量しか生成しない。その理由を明らかにするため本反応の解析を行ったところ,TVAIIは最終的な加水分解産物である,パノースによって強い阻害を受けることが明らかになり,このため,蓄積した糖転移生成物が分解されないで,安定に保持されるということを示した。 第3章は,両TVAの基質特異性の解析について述べている。TVAIはTVAIIより,高分子基質のデンプン,プルランに対してより優れた活性を持ち,逆にTVAIIは,オリゴ糖に対しより優れた活性を持つことを明らかにした。本実験の結果は,両酵素の性質はかなり異なっていることを示しており,TVAIは菌体外で,デンプンのような多糖の分解に関与していると推測され,TVAIIは,大腸菌のマルトース系の酵素であるMalZのように,菌体内でオリゴ糖の代謝に関与している酵素であることを示唆した。 第4章では,TVAIIの基質特異性の改変について述べている。TVAIとTVAIIの基質特異性の重要な違いに,パノースによる阻害があげられる。このパノースのTVAIIによる阻害に着目し,タカアミラーゼAの立体構造,および,TVA関連酵素をもとに,パノースの関与する残基を推測し,二種類の変異体を作成した。これらの変異体は,デンプン分解活性に対するプルラン分解活性の比が大幅に低下し,変異を導入した部位がプルラン分解活性に関与していることが示された。さらに,デンプン分解活性に対するシクロデキストリン分解活性の比は,一方の変異酵素では上昇したが,もう一方では低下し,変異を導入した部位によって異なる機能を持つことを示唆した。 第5章は総合討論である。以上の研究に討論を加え,本研究では糖転移産物の合成研究と遺伝子工学を組み合わせるという手法によって,酵素の分子認識機構の解明のみならず,新たなオリゴ糖合成法の開発という利用面についても多大な成果が得られたことを結論している。 以上本論文は,糖転移反応を利用したオリゴ糖の新たな合成法を開発し,本合成で得られた基質を用いてTVAの基質特異性の解析を行うことにより両アミラーゼの基質特異性の差を明確にし,さらに本結果をもとに作成した変異酵素によって基質特異性の変換に成功したものであり,学術上,応用上重要な,アミラーゼの分子認識機構の解析に多大な貢献を与えたものである。よって審査員一同は,申請者に博士(農学)の学位を授与してしかるべきものと判定した。 |
 -アミラーゼの基質特異性の解析と応用
-アミラーゼの基質特異性の解析と応用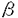 -ガラクトシド(IPTG)を添加することによって大量のアミラーゼを菌体内に生産することができる。pTNlには、lacプロモーターの下流にlac由来とTVAII遺伝子由来の二つのSDが存在し、両SDとも機能していることが分かった。しかし、このままでは、N末端が異なる二つのTVAIIが生産されてしまうので、lacSDまたはTVAIISDのみが働くように、欠失変異を導入したプラスミドを作成し、発現量の比較を行ったところ、TVAIISDで生産するプラスミドの方が、lacのそれよりも多いことが分かり、このプラスミドによって培地1lあたり100mg以上のTVAIIを得ることができた。
-ガラクトシド(IPTG)を添加することによって大量のアミラーゼを菌体内に生産することができる。pTNlには、lacプロモーターの下流にlac由来とTVAII遺伝子由来の二つのSDが存在し、両SDとも機能していることが分かった。しかし、このままでは、N末端が異なる二つのTVAIIが生産されてしまうので、lacSDまたはTVAIISDのみが働くように、欠失変異を導入したプラスミドを作成し、発現量の比較を行ったところ、TVAIISDで生産するプラスミドの方が、lacのそれよりも多いことが分かり、このプラスミドによって培地1lあたり100mg以上のTVAIIを得ることができた。