| 審査要旨 | | 本論文は大腸菌の細胞質膜の形成に関わると考えられるTolZ/FtsH蛋白質について解析したもので,三部から構成されている。生物一般において重要な役割を果たす膜系の機能発現機構の解明のため,精密な分子生物学的手法による解析が可能な大腸菌において,その細胞質膜の機能に関わる蛋白質の研究は必ず行われるべきものと考えられる。大腸菌のtolZ変異株は,コリシンE2,E3,D,Ia,Ibに対し耐性であり,グルコースでは生育できるが,非発酵性炭素源(コハク酸,ピルビン酸など)では生育できず,膜電位と電気化学的プロトン勾配の形成保持に欠陥をもつ。 著者は,この細胞質膜の状態の変化したtolZ変異株を用いてtolZ遺伝子のクローニングと解析を行い,更にtolZ変異株が非発酵性炭素源で生育できないという現象を解明するため,好気的呼吸鎖電子伝達系の解析を行った。 第一部では,tolZ遺伝子のクローニングを行った。著者は,大腸菌のcosmid DNAライブラリーを用い,tolZ変異を相補する40kbのcosmid DNAを分離した。次に,切り縮めたDNA断片からrecA-のtolZ変異株を部分的に相補する1.8kbのBamHI断片を取ることに成功した。この1.8kbのBamHI断片の全塩基配列の決定を行った結果,大腸菌ftsH遺伝子の配列と完全に一致したこと,更に,Hfr菌株を用いての接合の方法やP1形質導入などの方法でtolZ遺伝子が大腸菌染色体地図上69分に位置することが判明した。更に,ftsH遺伝子をもつプラスミドはtolZ変異を完全に相補することから,tolZ遺伝子はftsH遺伝子と同一のものであると結論した。 大腸菌FtsH蛋白質は,約71kDaの細胞質膜蛋白質で, ファージのcII蛋白質と転写因子 ファージのcII蛋白質と転写因子 32を特異的に分解するATPならびに亜鉛依存性のプロテアーゼであることが明らかにされている。現在まで,tolZ変異以外にも,std(stop transfer defective),hflB(high frequency of lysogenization)変異のいずれもftsH遺伝子の中に起こっ変異であることが相次いで判明した。ところが,tolZ変異株はStd-,Hfl-の表現型を示すが,ほかのftsH変異株のいずれもtolZ変異株の表現型を示さない。これらの表現型の違いを明かにするため,tolZ21変異遺伝子のクローニングとその塩基配列の決定を行った。その結果,亜鉛依存性プロテアーゼの活性中心モチーフHEXXHの二つ目のHis418がTyrに変化していた。tolZ変異株の表現型はこのHisの変異によってプロテアーゼの機能が失われたことによるものであることを発見した。 32を特異的に分解するATPならびに亜鉛依存性のプロテアーゼであることが明らかにされている。現在まで,tolZ変異以外にも,std(stop transfer defective),hflB(high frequency of lysogenization)変異のいずれもftsH遺伝子の中に起こっ変異であることが相次いで判明した。ところが,tolZ変異株はStd-,Hfl-の表現型を示すが,ほかのftsH変異株のいずれもtolZ変異株の表現型を示さない。これらの表現型の違いを明かにするため,tolZ21変異遺伝子のクローニングとその塩基配列の決定を行った。その結果,亜鉛依存性プロテアーゼの活性中心モチーフHEXXHの二つ目のHis418がTyrに変化していた。tolZ変異株の表現型はこのHisの変異によってプロテアーゼの機能が失われたことによるものであることを発見した。 第二部では,tolZ変異株が非発酵性炭素源で生育できないことから,好気的呼吸鎖電子伝達系成分に欠陥があると予想し,それらの成分を詳細に解析した。 野生型株及びtolZ変異株を30℃と40℃で培養し,対数増殖期,定常期の菌体から細胞質膜を調製した。各々の細胞質膜について,b型シトクロム,シトクロムo,シトクロムd含量を調べた。その結果,対数増殖期の40℃で,tolZ変異株のシトクロムo含量が減少していること,定常期では,温度によらずb型シトクロム,シトクロムo,シトクロムd含量のいずれも野生型株に比較して,tolZ変異株では少ないことが判明した。また,シトクロムb556をもつコハク酸脱水素酵素の活性は,野生型株に比較してtolZ変異株では少ない活性しか認められなかった。 更に,定常期の菌体から調製した細胞質膜の極低温での酸化還元差スペクトル,(還元型+一酸化炭素)-還元型の差スペクトルからtolZ変異株のシトクロムbd複合体に久陥があることを明らかにした。そして,それらの酵素を精製し,極低温での酸化還元差スペクトルの測定により,これらの欠陥を確認した。 以上の研究から,tolZ変異株において,呼吸鎖末端酸化酵素とコハク酸脱水素酵素のシトクロムに異常があることを明らかにした。tolZ変異株はこのような好気的呼吸鎖電子伝達系成分に欠陥があるために,非発酵性炭素源で生育できなくなると結論した。 これらの解析に基づき,TolZ/FtsH蛋白質は,シトクロムbo複合体,シトクロムbd複合体,コハク酸脱水素酵素のような膜内在性蛋白質の細胞質膜への組み込み,構造形成あるいは機能発現などに関与していることが示された。 第三部においては総合討論を行っている。 以上,本論文は大腸菌の細胞質膜の形成におけるTolZ/FtsH蛋白質の機能の一端を解明し,真核生物,原核生物を問わず最近次々と発見されているTolZ/FtsH蛋白質と相同性の高い他の蛋白質の研究にも大きな指針を与えるものと考えられる。したがって本論文は,学術上,応用上貢献するところが少なくない。よって審査員一同は申請者に博士(農学)の学位を授与してしかるべきものと判定した。 |
 -ラクタマーゼ前駆体のプロセシングやペニシリン結合蛋白質3の細胞質膜へのアセンブリー等に関与すること、また
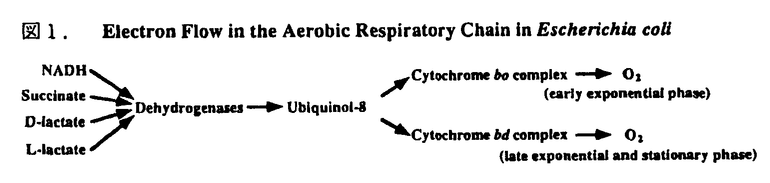
-ラクタマーゼ前駆体のプロセシングやペニシリン結合蛋白質3の細胞質膜へのアセンブリー等に関与すること、また 帯)を示すヘムbと、還元状態で一酸化炭素を結合する高スピンヘムoとCuBを補欠分子族として含み、本酵素の酸化還元反応中心として働いている。一方、シトクロムbd複合体は、二つのサブユニットからなっており、サブユミットIは極低温での酸化還元差スペクトルで556nmと559nmの吸収極大を示すヘムbとユビキノン結合部位を含み、サブユニットIIはヘムdをもっている。本酵素は、対数増殖期後期から定常期、相対的低い酸素分圧下で合成される緑色のシトクロムである。
帯)を示すヘムbと、還元状態で一酸化炭素を結合する高スピンヘムoとCuBを補欠分子族として含み、本酵素の酸化還元反応中心として働いている。一方、シトクロムbd複合体は、二つのサブユニットからなっており、サブユミットIは極低温での酸化還元差スペクトルで556nmと559nmの吸収極大を示すヘムbとユビキノン結合部位を含み、サブユニットIIはヘムdをもっている。本酵素は、対数増殖期後期から定常期、相対的低い酸素分圧下で合成される緑色のシトクロムである。