| 審査要旨 | | アクチンは筋細胞,非筋細胞を問わず全ての細胞に存在し,細胞骨格あるいは細胞運動の要のタンパク質である。これまでサイトカラシン類がアクチン重合阻害剤として広く研究に用いられ,各種細胞機能におけるアクチンの生理的役割が検討されてきた。本研究では,各種の海産毒の生理活性を検討する過程で,3つのマクロライド,すなわちミカロライドB(ML-B),アプリロニンA(AP-A),ビステオネライドA(BT-A)がアクチンを脱重合させることを見出した。生化学的な検討により,これらの海産毒の作用はサイトカラシン(CD)とは全く異なった機構であることが明らかとなった。論文は3章から構成され,第1章でアクチンに対する作用を生化学的に解明し,これを受けて,第2章では筋肉系のアタトミオシンに対する作用について,第3章では平滑筋組織に対する作用を明らかにしている。 第1章 アクチンに対する作用:CDはF-アクチンの成長端側を蓋って平衡を脱重合側へ変化させるため,比較的低濃度でアクチンの重台な抑制するが,核形成を促進し,過剰に投与しても完全には重合を抑制できない。これに対して3つのマクロライドはいずれもG-アクチンとのstoichiometryに従ってアクチンの重合を100%抑制した。また,3つの海産毒は核形成を促進しなかった。一方,4種類の物質は何れもF-アクチンを脱重合させた。この時,ML-Bと,AP-AはF-アクチンを切断したが,BT-A,CDでは切断作用は見られなかった。この様な成績から,これら3種のマクロライドは共にG-アクチンに結合して重合できないようにこれを隔離する作用を持つこと,ML-BとAP-Aはこれに加えてF-アクチンを構成するG-アクチンと結合してF-アクチンをその部分で切断するものと考えられた。このときML-BおよびAP-AはG-アクチンと1:1の複合体を形成し,BT-Aは1:2の複合体を形成することが示唆された。さらに結合実験および超遠心分析によってもBT-Aを介してアクチンが2量体になっている可能性が支持された。このことは3者には共通した構造があり,さらにBT-Aは2量体構造であるためこの共通した構造を2個もつことと一致した。 第2章 アクトミオシン系に対するミカロライドBの抑制作用:ミオシンはATPase活性を持ち,F-アクチンによってその活性が増強される。このアクチンとミオシンの相互作用が細胞運動の原動力となる。ウサギ骨格筋より抽出したアクチンおよびミオシンによって再構成したアクトミオシン系のATPase活性をCDはほとんど抑制しなかった。他方,ML-Bはミオシン単独のATPase活性を全く抑制しなかったものの,F-アクチンによって増強された分のみを抑制した。ミオシン頭部(SI)のATPase活性もF-アクチンによって増強されるが,ML-BはこのアタトSIのATPase活性に対してもアクチンとの濃度比が1以上の時に最大抑制を示した。したがってML-BがアクトミオシンおよびアクトSIのATPase活性を抑制するのは,F-アクチンがML-Bによって脱重合され,ATPase活性を増強できなくなったためと考えられた。 第3章 平滑筋収縮に対する抑制作用:平滑筋は外部からの刺激によって細胞内のCa濃度が上昇すると20kDaのミオシン軽鎖がリン酸化されて収縮する。CDおよびML-B,BT-A,AP-Aはいずれもラット大動脈平滑筋の細胞内Ca濃度は変化させずに収縮のみを抑制した。また脱膜化標本の収縮をML-BとCDはともに抑制した。以上の結果からML-BとCDは平滑筋の収縮蛋白系のうちアクチンフィラメントに作用して収縮を抑制するものと考えられた。またAP-AおよびBT-Aも同様の作用を持つことが示唆された。さらにCDもこの収縮を100%抑制することから,収縮な抑制するためには必ずしもフィラメントが完全に破壊される必要はないことが示唆された。 以上を要約すると,ML-B,AP-A,BT-Aの3つの海産毒がCDとは異なった機序によってアクチンの重合を阻害することを明らかにした。さらに平滑筋組織を用いた実験からML-Bはアクチンに対する選択性が高いことが示され,これらの物質が今後アクチン機能に対するプロープとして広く使われることが期待された。この様に,本研究は学術上貢献するところが少なくない。よって審査員一同は本論文が博士(獣医学)の学位論文として価値あるものと認めた。 |
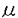 Mであった。BT-Aのstoichiometryはさらに結合実験および超遠心分析によっても確認した。精製したG-アクチンにBT-Aを結合させ、限外濾過膜で上清の遊離BT-Aを分離し、濃度をHPLCで測定した。その結果、BT-Aはアクチンとの結合において正の協同性を持つと推測され、結合部位数が1.7、Kdは0.15
Mであった。BT-Aのstoichiometryはさらに結合実験および超遠心分析によっても確認した。精製したG-アクチンにBT-Aを結合させ、限外濾過膜で上清の遊離BT-Aを分離し、濃度をHPLCで測定した。その結果、BT-Aはアクチンとの結合において正の協同性を持つと推測され、結合部位数が1.7、Kdは0.15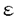 ATP)の蛍光を指標としてアクチンがATPを交換する速度を測定した。10分間G-アクチンをBT-Aで処置すると
ATP)の蛍光を指標としてアクチンがATPを交換する速度を測定した。10分間G-アクチンをBT-Aで処置すると