本研究は、多細胞生物の発生において重要な細胞の増殖と分化の制御を検討するために、筋細胞を用いてその増殖と分化を制御する細胞外因子の探索とその細胞内機構を解析し、下記の結果を得ている。 1.マウス樹立筋芽細胞株C2C12の増殖分化を制御する細胞外因子を無血清分化誘導条件下で検索した結果、リゾフォスファチジン酸(LPA)が、bFGFと同様、C2C12細胞に対し増殖促進・分化抑制的に作用することを見いだした。これらの作用はいずれも数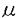 Mの濃度で引き起こされ、以前より報告のあった線維芽細胞の増殖促進能とほぼ同様の濃度で作用した。更にLPAと構造的に類似している種々のリン脂質は同様の活性を示さないことから、これらの作用がLPAにより特異的にもたらされていることが明かとなった。 Mの濃度で引き起こされ、以前より報告のあった線維芽細胞の増殖促進能とほぼ同様の濃度で作用した。更にLPAと構造的に類似している種々のリン脂質は同様の活性を示さないことから、これらの作用がLPAにより特異的にもたらされていることが明かとなった。 2.LPAのC2C12細胞に対する増殖促進・分化抑制活性は共に百日咳毒素により阻害され、Giタンパク質を介することが明かとなった。一方、bFGFの活性は阻害されず、Giの関与は否定された。LPAを新しく同定したことにより、その存在は示唆されていたものの解析のなされていなかったGiを介する増殖分化制御機構の解析が可能となった。また、LPAとbFGFそれぞれ十分量共存させるとその増殖促進能および分化抑制能に相乗効果が認められ、LPAとbFGFが異なった細胞内機構を用いて作用していることが示唆された。 3.LPAとbFGFの、筋分化において中心的な役割を演ずるMyoDファミリーの筋分化因子(MyoD,myogenin,Myf-5,MRF-4)に対する制御を検討した結果、LPAとbFGFが以下のようにMyoDファミリーの筋分化因子に対して異なった制御を行っていることが明かとなった。(1)LPAがMyoDの発現を維持しつつ筋芽細胞の増殖を維持しているのに対して、bFGFはMyoD自身の発現を抑制していた。この時タンパク質、mRNA共に減少し、転写レベルでの制御を受けていた。(2)MyoDとは対照的に,LPA,bFGF共にmyf-5のmRNA発現には大きな影響を与えていなかった。(3)LPA,bFGF共にmyogeninの発現はmRNA,蛋白質レベル共に抑制していた。(MRF-4はこの系では検出限界以下であった。) 4.LPAとbFGFによる増殖促進・分化抑制活性には細胞レベルでも明確な差異が認められた。C2C12筋芽細胞をLPA,bFGFまたは血清の存在下で長期間(5日)培養しコンフルエントに達すると、LPAで培養した場合は分化が誘導され、myogeninと共に最終分化のマーカーが発現し筋管を形成した。これに対して、bFGFで培養した場合は分化は強力に抑制され、筋管細胞はほとんど形成されなかった(このbFGFの効果はLPAとの共存により増強された)。このときはmyogeninのみならずMyoDの発現も抑制されたままであり、bFGFの作用は細胞同志の接触よりも優位であると考えられた。 5.以上のbFGFと異なるLPAの性質は、基本的に血清と共通であり、LPAが血清中に1-5M含まれていることを考慮すると、血清の筋芽細胞に対する増殖促進・分化抑性能の多くをLPAが担っている可能性が示された。 以上のように、本論文はLPAを新たな筋芽細胞増殖促進、分化抑制因子として同定し、LPAがbFGFとは異なる細胞内機序により筋芽細胞の増殖と分化を制御していることを明かにした。本研究は筋細胞の増殖と分化の制御を検討するうえで重要な知見を与えるものであり、また、他の細胞系では明かとはならなかったチロシンキナーゼ系とG蛋白質系の増殖刺激シグナルの作用点の差異を明確にした点でもその意義は大きいと考えられる。以上のことから、本研究は学位の授与に値するものと考えられる。 |