異物に対する生体の免疫反応は細胞性免疫と液性免疫に大別され、それぞれ異なるヘルパーT細胞サブセットから産生される一群のリンホカインによって調節されている。即ち、Th1細胞はIL-2、IFN-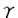 、lymphotoxinなどを産生しマクロファージ等を活性化するのに対し、Th2細胞はIL-4、IL-5、IL-10などを産生しB細胞の抗体産生を促進する。しかし、これらの細胞による異なるリンホカイン発現機構の詳細は不明である。本研究ではこのサブタイプ特異的なリンホカイン発現の調節機構を明らかにするために、主にTh2細胞から産生されるIL-5の遺伝子発現機構の解析をTh1特異的リンホカインであるIL-2と比較しながら行った。 、lymphotoxinなどを産生しマクロファージ等を活性化するのに対し、Th2細胞はIL-4、IL-5、IL-10などを産生しB細胞の抗体産生を促進する。しかし、これらの細胞による異なるリンホカイン発現機構の詳細は不明である。本研究ではこのサブタイプ特異的なリンホカイン発現の調節機構を明らかにするために、主にTh2細胞から産生されるIL-5の遺伝子発現機構の解析をTh1特異的リンホカインであるIL-2と比較しながら行った。 IL-5遺伝子はヒト5番染色体、マウス11番染色体上にIL-3,IL-4,GM-CSF,IL-13といった一群のリンホカイン遺伝子とクラスターをなして存在している。また、IL-5遺伝子のプロモーター領域は他のリンホカイン遺伝子にも共通して見いだされる配列(CLE0,CLE1,CLE2)を含んでいる。 IL-2遺伝子の発現調節機構は多くのグループにより研究され、その主な調節領域はプロモーターの上流300bpの内に存在することが示されている。一方、IL-5遺伝子の発現調節機構に関しては、株化されたT細胞においてフォルボールエステル(PMA)やカルシウムイオノフォア(A23187)刺激により誘導されるIL-5の産生レベルは低く、さらに、IL-5遺伝子のプロモーターは種々のT細胞における一過性発現系においてこれらの刺激に応答しなかったため、その転写調節機構の解析がなされていなかった。 そこで本研究では(1)まずIL-5を産生する細胞とそのための最適なシグナルの検索を行い、(2)株化T細胞における一過性発現系を樹立し、(3)IL-5プロモーター上のcis-regulatory elementおよびこれに結合する因子を同定、(4)これから得られた結果をTh1/Th2細胞を用いて検討するといったアプローチを取った。 IL-5遺伝子発現におけるcAMPシグナルの効果 一般に、T細胞の活性化シグナルはPMAとA23187により代替され、株化されたT細胞やT細胞クローンではこれらの刺激によりリンホカインの産生が誘導される。最近、T細胞の活性化にはT細胞レセプターだけではなくCD28、LFA1といった膜分子からのシグナルも重要であること、また、各々のリンホカインの産生が誘導されるための最適のシグナルはそれぞれ異なるということが明らかになってきた。また、PGE2など細胞内のcAMPレベルを上げる物質はTh1細胞のリンホカインの産生は阻害するが、Th2細胞のリンホカイン産生は阻害せず、特にIL-5の産生は増加させることが分かってきた。 ELISAおよびNorthern blotによりマウス胸腺腫EL-4細胞においてIL-5発現に必要なシグナルの解析を行ったところ、PMA単独の刺激では低レベルのIL-5の産生しか見られないが、PMAとcAMPの両方の刺激を加えることによりタンパク質およびmRNAレベルにおいて顕著な産生の増大が見られた。逆にcAMPはPMA依存的なIL-2の産生をほぼ完全に阻害した。また、IL-5(5’上流領域約1200bp)とIL-2(5’上流領域約500bp)のプロモーターをそれぞれルシフェラーゼ遺伝子に連結し、EL-4細胞で一過性発現させたところ、各々のシグナルに対するプロモーターの活性は内在性の遺伝子の応答性と一致した。さらに、プロテインキナーゼA(PKA)の触媒サブユニットcDNAのコトランスフェクションによりcAMP添加と同様な効果が見られた。以上の結果から、PKAのシグナル伝達経路はIL-5遺伝子発現には促進的に、またIL-2の遺伝子発現には抑制的に働くことが明らかになった。 IL-5遺伝子プロモーターの解析 次に、PMAとcAMPシグナルに反応する領域を同定するために、IL-5プロモーター5’上流領域の様々な欠失変異体や突然変異体を作製し、EL-4細胞において一過性発現させてこれらのシグナルに対する反応性を調べた。その結果、IL-5A(-948--932),IL-5P(-117--91),IL-5C(-74--56),IL-5CLE0(-55--38)の4つのelementが、PMAとcAMPシグナルによるIL-5プロモーターの活性化に必要なelementとして同定された。これらの配列の中にはCRE様の配列は認められないため、新たなcAMPの作用機構が予想される。また、IL-5P,IL-5C,IL-5CLE0の配列はヒトのIL-5遺伝子においても非常によく保存されており、ヒトのIL-5プロモーターを用いた解析でも同様の結果が得られた。 IL-5PおよびIL-5CLE0に結合する因子の解析 これらのelementの配列を注意深く観察したところ、IL-5PとIL-5CLE0の配列がIL-2プロモーターのNF-AT配列と類似していることから、この2つのelementに関してさらに解析を行った。EL-4細胞の核抽出液を用いたゲルシフトアッセイの結果、IL-5P配列に結合する複合体(NFIL-5P)が検出された。NFIL-5Pは(1)刺激により誘導される(2)転写因子NF-ATと同様な細胞分布を示す(3)NF-AT配列により競合される(4)免疫抑制剤のcyclosporin Aおよび蛋白合成阻害剤のcyclohexamideにより抑制される(5)COS-7細胞に発現させたrecombinant NF-ATと転写因子AP1により再構成されるなどのことから、転写因子NF-ATと類似した因子でであると結論した。 IL-5CLE0には構成的(aとc)及び誘導的複合体(b)が結合する。 NF-ATやIL-5Pの配列との競合実験から、これらの複合体はNF-ATとは構成成分を共有するが、異なる因子であることを示した。以上の結果から、IL-5とIL-2遺伝子はcAMPにより相反的に調節されるにも関わらず、ともにNF-AT様の配列により制御されることが明らかになった。 Th1およびTh2細胞におけるIL-5遺伝子の発現 以上に示したように、Th0細胞のリンホカイン産生パターンを示しているEL-4細胞において、cAMPシグナルはIL-2遺伝子のプロモーターには抑制的に、IL-5遺伝子のプロモーターには促進的に働く。IL-2とIL-5はそれぞれTh1とTh2細胞から特異的に産生されることから、EL-4細胞で見られるcAMPのIL-2遺伝子およびIL-5遺伝子のプロモーターに対する相反的効果はTh1/Th2細胞における特異的リンホカイン発現の調節機構と関連があると予測した。実際に、IL-2遺伝子のプロモーターとIL-5遺伝子のプロモーターをHDK1(Th1)およびD10(Th2)細胞に導入した結果、IL-2遺伝子のプロモーターはTh1細胞のみで、IL-5遺伝子のプロモーターはTh2細胞のみで活性が認められた。また、複数コピーのIL-5CLE0配列はSV40の基本プロモーターをD10細胞では活性化するが、HDK1細胞では活性化しないことから、IL-5CLE0配列がTh2細胞特異的なIL-5遺伝子発現にかかわる可能性が示唆された。。 以上、本研究ではEL-4細胞においてIL-5の遺伝子発現にPMAとcAMPの両シグナルが関与していることを証明し、これらのシグナルに反応する5’上流領域上の4つの機能的cis-element(IL-5A,IL-5P,IL-5C,IL-5CLE0)を同定した。特に、IL-5Pを介したプロモーターの活性の調節にはNF-AT様の因子(NFIL-5P)が関与しており、また、これと類似配列を持つIL-5CLE0にはNF-ATとは異なる構成的及び誘導的因子が結合することを見い出した。また、IL-2およびIL-5遺伝子のプロモーターがそれぞれTh1およびTh2細胞において特異的に活性化され、IL-5遺伝子のTh2細胞における特異的に発現にIL-5CLE0が関与することを示した。今後、cAMPシグナルのT細胞活性化における生理的な意義及びTh1/Th2細胞における特異的リンホカイン遺伝子発現への関与の可能性を明らかにすることが課題である。また、本研究で同定された4つのcis-elementのTh1とTh2細胞における機能、また、Th2細胞においてIL-5CLE0の機能がどのように制御されるかを明らかにすることより、Th1とTh2細胞におけるリンホカイン遺伝子の協調的及び特異的発現の調節機構を説明する手掛かりが得られるものと期待される。 |