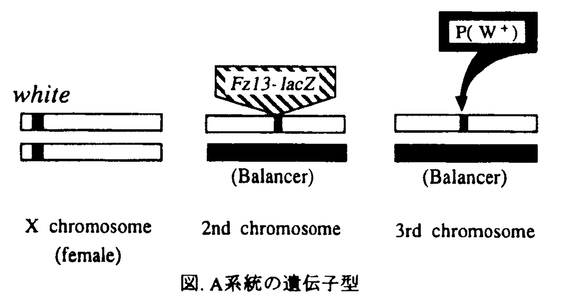
本研究はショウジョウバエの幼虫期神経発生機構の解明のため、P因子を用いた突然変異体の作成をおこない、表現型の解析及び突然変異遺伝子の分子生物学的解析をおこなったものであり、下記の結果を得ている。 1.幼虫期神経芽細胞でlacZの発現が見られるエンハンサートラップ系統Fz13をマーカーとして用い、そのlacZの発現に異常を示すP因子挿入突然変異体をスクリーニングした。この結果、約700系統の中から幼虫期/蛹期劣勢致死突然変異A9-Fz13系統を得た。この系統は幼虫期以降の神経系に顕著な異常を示し、特に成虫型神経系に見られるoptic lobeが全く形成されていないことがわかった。この系統では未同定の遺伝子座にP因子挿入突然変異が導入されたことがで明らかとなったため、上記の表現型よりこの遺伝子座をno optic lobes(nol)と名付けた。 2.ショウジョウバエの神経細胞を特異的に認識する抗HRP抗体を用いて、nolの中枢神経系を観察した結果、胚期の神経発生には顕著な異常が見られず幼虫期以降の神経発生に特異的に異常を示すことが明らかとなった。 また、BrdU(ブロモデオキシウリジン;チミジンのアナログ)を用い幼虫期神経芽細胞の増殖パターンを観察したところ、幼虫初期のnolでは一部の例外を除き神経芽細胞へのBrdUの取り込みが全く見られず、幼虫期神経芽細胞の再分化が正常におこなわれていないことが明らかとなった。一方、幼虫後期になると、nolでは神経芽細胞とは明らかに異なる位置に分布する細胞でBrdUを取り込んだ細胞が見られる。また、抗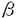 -チューブリン抗体を用いて観察したところ、この時期の中枢神経系では分裂期の微小管がほとんど観察されないことから、これらの細胞はほとんど分裂せず倍数体となっていることが示唆された。 -チューブリン抗体を用いて観察したところ、この時期の中枢神経系では分裂期の微小管がほとんど観察されないことから、これらの細胞はほとんど分裂せず倍数体となっていることが示唆された。 3.nol遺伝子を分子生物学的に解析するため、下記のようにこの遺伝子のクローニングを試みた。すなわち、P因子をプローブとしてA9系統のゲノムDNAをクローニングした結果、P因子挿入位置近傍約20kbにわたるゲノムDNA断片を得ることができた。これらのゲノムDNA断片をプローブとして用いることによりノーザンブロットをおこなった結果、この領域には3つの転写単位、transcript A、B、Cがあることがわかった。nolホモ接合体のmRNAを用いたノーザンブロットから、このうちtranscript Aはnolホモ接合体で顕著に発現量が減少していることが明らかとなった。また、in situハイブリダイゼーションの結果、この転写単位が幼虫期中枢神経系で発現が見られることがわかった。 これらの結果から、transcript Aがnol遺伝子であることが示唆されたためcDNAのクローニングを行い、この転写単位の塩基配列を決定した。この結果、transcript Aからは130アミノ酸のポリペプチド鎖が翻訳されることが推定され、そのアミノ酸配列からその遺伝子産物は分泌性の糖タンパク質であることが示唆された。 以上本論文は、P因子の導入による突然変異体の作成及びその解析から、ショウジョウバエの幼虫期神経発生に関与する新たな遺伝子を同定し、その遺伝子産物と推測されるタンパク質を同定したものである。本研究は、初期神経発生機構の解明に重要な貢献をなす可能性を蔵しており、学位の授与に値するものと認められる。 |