学位論文要旨
| No | 111345 | |
| 著者(漢字) | 田中,晃一 | |
| 著者(英字) | ||
| 著者(カナ) | タナカ,コウイチ | |
| 標題(和) | 分裂酵母Schizosaccharomyces pombe のG1/S期移行を制御する新規細胞周期調節遺伝子の発見と解析 | |
| 標題(洋) | Identification and characterization of novel cdc genes that regulate G1/S transition in fission yeast Schizosaccharomyces pombe | |
| 報告番号 | 111345 | |
| 報告番号 | 甲11345 | |
| 学位授与日 | 1995.03.29 | |
| 学位種別 | 課程博士 | |
| 学位種類 | 博士(医学) | |
| 学位記番号 | 博医第999号 | |
| 研究科 | 医学系研究科 | |
| 専攻 | 第二基礎医学専攻 | |
| 論文審査委員 | ||
| 内容要旨 | 真核細胞の細胞周期は主にG1期とG2期で調節される。G2期の役割がS期の完了を確認し、M期を開始させることにあるのに対し、G1期の役割は外部環境の変化に反応して増殖の進行あるいは停止、もしくは分化への移行を決定することにある。この決定は絶対的なもので、一度G1期からS期へ進行してしまうとそのサイクルが終了するまで分化過程に移行することはできない。このような遷移点は酵母ではstart、動物細胞ではrestriction point とよばれ、古くからその存在が知られていた。G1/S期制御機構の解明は細胞周期だけでなく、細胞の不死化、癌化、あるいは細胞死といった現象を理解するために必要不可欠である。しかし、制御が複雑な高等動物細胞を用いた解析は非常に困難であることが多い。近年、細胞周期の基本的な調節機構が酵母などの下等真核生物からヒトを含む高等真核生物まで進化的に高度に保存されていることが示されたため、私は遺伝学的な解析が容易な分裂酵母をモデルとして、真核細胞に普遍的に存在する基本的なG1/S期制御機構を明らかにすることを試みた。 G1後期からS期にかけて種々の遺伝子の発現が一過的におこる現象は酵母からヒトに至るまで多くの生物種において見いだされる。出芽酵母では、このような細胞周期特異的な遺伝子発現は主にSCB,MCBとよばれる2種のシス配列により制御されている(図)。SCBはG1サイクリン遺伝子など、MCBはDNA複製に関与する酵素をコードする遺伝子などのプロモーター領域に存在し、それぞれSwi4及びMbp1というDNA結合タンパク質により特異的に認識される。両DNA結合タンパク質はSwi6とよばれる共通の調節サブユニットタンパク質とヘテロ二量体を形成し、G1/S期特異的な転写因子複合体として機能する。興味深いことにMCBと全く同じシス配列を介した細胞周期特異的な遺伝子発現制御が、進化的に遠く隔たった分裂酵母においても見いだされている。リボヌクレオチド還元酵素をコードするcdc22+遺伝子もその一種で、プロモーター領域に数カ所のMCBを有し、やはりG1後期からS期にかけて一過的に発現が上昇する。最近、分裂酵母のCdc10タンパク質がcdc22+のMCBに結合する転写因子複合体の構成因子であることが報告された。Cdc10はstartの制御に必須の機能を持ち、出芽酵母のSwi4,Swi6,Mbp1と相同性を示すことが知られている。これら全てのタンパク質は、タンパク質間の相互作用に関与すると考えられているcdc10/SWI6/ankylinモチーフを中央部に二組持つのが特徴である。 私は過剰発現により、cdc10+遺伝子の温度感受性突然変異株(cdc10ts)を非常に強く相補できるres1+遺伝子を単離した。res1+遺伝子産物はやはり中央部に二組のcdc10/SWI6/ankylinモチーフを持っていたが、それに加えてN末端領域でSwi4,Mbp1と顕著な相同性を示した。Swi4,Mbp1のN末端領域は、SCB及びMCBシス領域を特異的に認識するDNA結合部位であることが知られている。一方、Swi6のC末端領域はSwi4,Mbp1との結合部位であるが、Cdc10のC末端領域と構造が類似している。従って構造的にはRes1はSwi4,Mbp1といったDNA結合サブユニットに、Cdc10はSwi6調節サブユニットに対応していると考えられた。次にres1+遺伝子の機能を探るため、その遺伝子破壊株( Res1は直接シス配列を認識するDNA結合タンパク質であり、Cdc10単独では転写因子として機能できないと考えられる。しかしながら、res1+遺伝子を破壊してもその株は完全に致死とはならない。その理由としてres1+と重複した機能を持つ別の遺伝子の存在が予想されたため、 Res1,2-Cdc10に関与する因子を同定する目的でcdc10tsを相補できる遺伝子をスクリーニングし、rep1+遺伝子を単離した。rep1+はC末端にZnフィンガー構造を有するタンパク質をコードし、過剰発現により 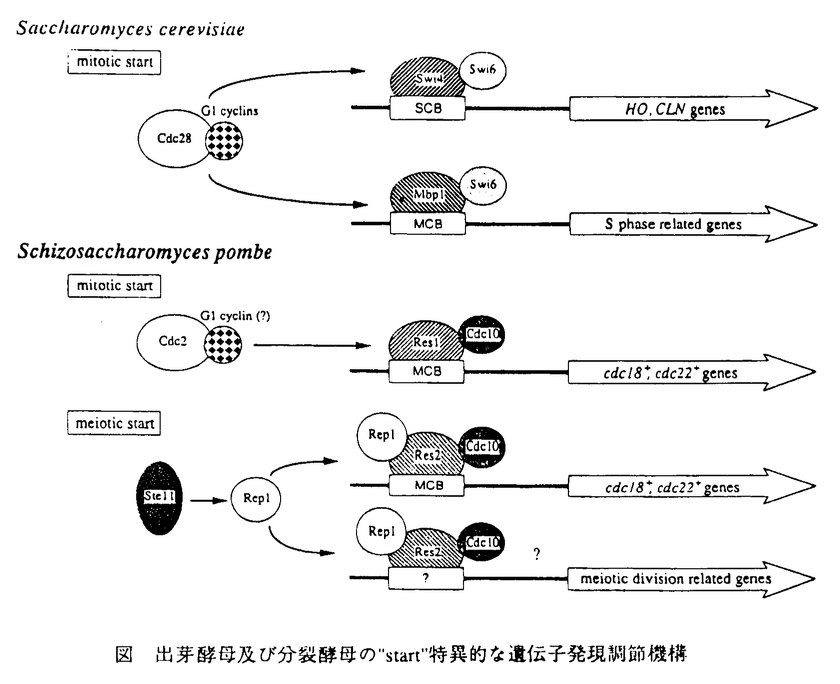 | |
| 審査要旨 | 本研究は真核生物の細胞周期調節機構を明らかにするため、モデル系として分裂酵母を材料とし、そのG1/S期調節に関わる遺伝子群の単離・解析を行ったものであり、下記の結果を得ている。 1.cdc10+遺伝子はG1期に特異的な転写調節因子複合体の構成因子をコードする。その過剰発現により、cdc10+遺伝子の温度感受性突然変異株を非常に強く抑圧できるres1+遺伝子を単離した。res1+遺伝子産物はそのN末領域において出芽酵母のSwi4蛋白質のDNA結合領域と高い相同性を示した。欠失変異解析によりこの領域はRes1蛋白質の機能に必須であることが明らかとなった。 2.res1+遺伝子を破壊した株は生存率が著しく低下した。かろうじて生育できたものも高温および低温では増殖を停止し、典型的なcdc表現型を示した。FACS解析および増殖を停止した状態での接合実験の結果より、res1+遺伝子の機能はG1期のSTART制御に必要であることが示された。 3.res1+遺伝子はcdc10+遺伝子破壊株の致死性をも相補することができた。しかし、cdc10+遺伝子はres1遺伝子破壊株の高温および低温感受性を相輔できなかった。以上の構造的・機能的な特徴より、Res1蛋白質はDNA結合サブユニットとして、Cdc10蛋白質は調節サブユニットとして、G1期に特異的な転写因子複合体を構成することが示唆された。 4.res1遺伝子破壊株の低温感受性を抑圧できる活性を指標としてres2+遺伝子を単離した。res2+遺伝子産物はRes1蛋白質と最も高い相同性を示した。res2+遺伝子破壊株は体細胞分裂過程において特に顕著な欠損を示さなかった。しかし、res1+遺伝子とres2+遺伝子を同時に破壊した株は完全に致死となったことより、体細胞分裂過程においてres1+遺伝子とres2+遺伝子は重複した機能を有することが明らかとなった。 5.res2+遺伝子の発現は減数分裂過程で強く誘導された。遺伝子破壊株の表現型の解析により、res2+遺伝子の機能は減数分裂前DNA合成の開始と、その後の核分裂の制御に関わることが示された。以上の結果より、分裂酵母のG1期に特異的な遺伝子発現制御にはRes1とRes2という良く似た蛋白質が重複して機能しており、前者は主に体細胞分裂過程、後者は主に減数分裂過程でより重要な役割を果たしていることが明らかとなった。 6.温度感受性cdc10変異株を宿主としてrep1+遺伝子を単離した。rep1+遺伝子はres1+遺伝子破壊株の高温および低温感受性も抑圧する活性を示した。rep1+遺伝子産物は他の蛋白質と特に顕著な相同性を示さなかったが、C末に1つのZnフィンガーモチーフを持っていた。 7.rep1+遺伝子破壊株の体細胞分裂過程は正常であった。しかし、一度G1期で増殖を停止させた後に減数分裂を誘導すると、rep1+遺伝子破壊株はres2+遺伝子の発現を誘導できず、減数分裂前DNA合成を開始できなかった。一方、栄養増殖からただちに減数分裂を誘導すると、rep1+遺伝子破壊株は減数分裂前DNA合成後の核分裂過程に異常を示した。しかし、rep1+遺伝子はres2+遺伝子の転写活性化を介することなしにcdc10変異株およびres1+遺伝子破壊株を抑圧できた。以上の結果よりrep1+遺伝子は減数分裂期特異的なres2+遺伝子の転写誘導および蛋白質レベルでのRes2の活性化により、減数分裂過程の制御に関わることが示唆された。 8.rep1+遺伝子の発現は体細胞分裂過程においてはほとんど見いだされず、窒素源の枯渇と接合フェロモン刺激により減数分裂過程に特異的に誘導されることが明らかとなった。 以上、本論文においては分裂酵母の体細胞分裂および減数分裂のSTART制御に働く2組の転写因子複合体(Res1-Cdc10,Res2-Cdc10)の存在を明らかにし、さらに減数分裂過程でRes2-Cdc10複合体の活性化因子として機能するRep1の同定を行った。本研究はG1期に特異的な転写調節機構を明らかにし、これまでほとんど未知であった分裂酵母のSTART制御機構に対する理解を深めた。また、基本的な細胞周期制御機構は進化を通じて保存されていると考えられることより、高等動物のG1/S期調節機構の解明にも重要な貢献をなすと考えられ、学位の授与に値するものと考えられる。 | |
| UTokyo Repositoryリンク | http://hdl.handle.net/2261/53856 |
 res1)を作成した。その結果、res1+遺伝子を破壊された細胞は生育が非常に悪くなり、高温及び低温ではstartを越えてS期に進行することができず、増殖をG1期で停止するという表現型を示した。また、破壊されたres1+遺伝子をホモに持つ二倍体細胞は減数分裂前DNA合成をほとんど開始することができなかった。cdc10ts株も同様に、制限温度下では増殖をstart前のG1期で停止し、その二倍体細胞は減数分裂前DNA合成を開始できない。これら両変異株の極めて類似した表現型は、Res1,Cdc10がG1期制御機構の中で同じ作用点を持つことを示唆している。更に、res1+遺伝子が
res1)を作成した。その結果、res1+遺伝子を破壊された細胞は生育が非常に悪くなり、高温及び低温ではstartを越えてS期に進行することができず、増殖をG1期で停止するという表現型を示した。また、破壊されたres1+遺伝子をホモに持つ二倍体細胞は減数分裂前DNA合成をほとんど開始することができなかった。cdc10ts株も同様に、制限温度下では増殖をstart前のG1期で停止し、その二倍体細胞は減数分裂前DNA合成を開始できない。これら両変異株の極めて類似した表現型は、Res1,Cdc10がG1期制御機構の中で同じ作用点を持つことを示唆している。更に、res1+遺伝子が