本研究は抗原提示細胞内で合成される、いわゆる内因性抗原蛋白のMHCクラスI分子を介する抗原提示の機構をより明らかにするために、抗原蛋白が細胞質あるいはミトコンドリアに局在した場合の抗原提示の解析を試みたもので、下記の結果を得ている。 1、抗原蛋白として用いた緑膿菌分岐アミノ鎖結合蛋白(Pseudomonas aeruginosa PAO Leu-Ile-Val-Binding Protein,以下BraC)のcDNAのみを組み込んだpSV1112BraCとBraCのcDNAの5’側にミトコンドリア移行シグナルを結合させたpSVmtBraC、BraCの5’側に核移行シグナルに続いてミトコンドリア移行シグナルを結合したpSVt-mtBraCの3種類を構築し、Balb c/3T3細胞株に導入し、1)細胞質のみにBraCが局在しているBC-15 2)ミトコンドリアのみにBraCが局在しているYZ-710 3)ミトコンドリア移行シグナルペプチドを持ちながら、核にBraCが局在するTM-35の3種類のトランスフェクタント細胞株を樹立した。 2、BraCを内在するそれぞれの細胞株をBALB/cマウスに免疫することによる細胞障害性T細胞(CTL)の誘導を試みた。BC-15により誘導されたCTLはBC-15に特異的で、YZ-710を障害しない。一方、YZ-710により誘導されたCTLはYZ-710に特異的でBC-15もTM-35も障害しなかったことから、抗YZ-710CTLはYZ-710細胞が提示するBraC由来ペプチドを認識すると考えられ、ミトコンドリア移行シグナル部分を認識しているのではない。 3、これらのCTLはCD8陽性で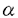 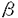 T細胞であった。さらにこれらのCTLによる細胞障害は古典的MHCクラスI拘束性である。 T細胞であった。さらにこれらのCTLによる細胞障害は古典的MHCクラスI拘束性である。 4、これらのCTLは共にアルカリ分解したBraCペプチドの存在下で親株Balbc/3T3を障害する。すなわち、これらの2種類のCTLは共にBraCそのものの断片ペプチドを認識する。さらにCNBr分解BraCの存在下で抗YZ-710CTLはBalb c/3T3を障害するが、抗BC-15CTLは障害しない。さらに抗YZ-710CTLの認識するエピトープペプチドがGYKLIFRTIであることを示した。これらの結果から、この2種類のCTLは認識するエピトープが異なることが示された。 以上、本論文は細胞質局在抗原のみならず、ミトコンドリアに局在する蛋白抗原もCD8T細胞に提示されること、そしてミトコンドリア局在抗原は細胞質局在抗原とは異なった抗原ペプチドを提示しうることを示したものであり、内因性抗原の提示機構の解明に資するものと考えられ、学位の授与に値するものと考えられる。 |