Ephはerythropoietin-producing human hepatocellular carcimoma cell line(ETL1)よりv-fpsをプローブとして単離されたレセプター型チロシンキナーゼである。 Ephの基本構造は、細胞膜外にシステインが多くみられる部位とファイブロネクチンIII型を2回繰り返す部位があり、細胞内にチロシンキナーゼ部位の構造をとり、これまで知られているレセプター型チロシンキナーゼの4型(Ullrich&Schlessingerが提唱)とは異なる独立したレセプター型チロシンキナーゼと考えられる。 近年Ephファミリーは次々と単離されている。Ephファミリーの多くは神経系で発現を認め、さらに顎下部増殖や神経前駆体に一過性発現をみることより、神経系発達に関与していることが示唆されている。しかしその機能は明らかでなく、またごく最近ElkおよびEckのリガンドが同定された以外、これまでEphファミリーのリガンドは同定されていない。 私はEphの細胞内情報伝達機構および生物学的活性を解析するために、EGFレセプター(EGFR)を用い、EGFRの細胞膜外領域およびEphの細胞膜貫通領域、細胞質内領域を融合したEGFR-Ephキメラレセプター(EGPR)cDNAを構築し、内因性EGFRを発現していない細胞株NIH3T3subline(3T3sub)(後藤典子博士東大医科研より供与)にEGPRcDNAとEGFRcDNAをリン酸カルシウム法でそれぞれトランスフェクトし、EGPRを高発現した安定変異株(EGP1)およびEGFRを高発現した安定変異株(EGF1)を作成し、解析を行った。またEphのATP binding site(Lys664→Met)を変異させたEphK664を作成し、EGPRの構築と同様の方法でEGFRとEphK664をキメラ結合させたEGPRK664を構築し、これを安定発現する細胞株(K664)を作成し解析に用いた。 さらにEphトランスジェニックマウスを作成し、Ephの生物学的活性を検討した。 1Ephポリクローナル抗体作成 グルタチオン-S-トランスフェラーゼ(GST)とEphのカルボキシル末端との融合蛋白質およびGSTとEphのアミノ末端との融合蛋白質を作成し、それらをウサギに免疫することで、抗カルボキシル末端(抗C)Ephポリクロナール抗体と抗アミノ末端(抗N)Ephポリクローナル抗体を作成した。 抗EGFRモノクローナル抗体(Oncogen sciense社)および抗C-Ephポリクローナル抗体を用い、EGPRがEGP1で、またEGFRがEGF1で安定高発現していることを確認した。 2EGF刺激によるチロシンリン酸化 細胞株EGP1、EGF1、およびK664をEGF刺激し、細胞内におけるレセプター自身のチロシンリン酸化およびある種の細胞内タンパク質のリン酸化を確認した。EGF1ではEGF刺激後に約50、65、85、120、170、190kDa付近にチロシンリン酸化バンドを認めた。これまでの報告より、50kDaはShc、65kDaはp62(GAP associated protein)、85kDaはphosphatidylinositol 3’kinase(PI3kinase)、120kDaはGAP、170kDaはEGFR自身、190kDaはp190(GAP associated protein)のチロシンリン酸化と推定された。EGF刺激後のEGP1においては、約65、120、150、190kDaにチロシンリン酸化バンドが確認され、p150はEGPR自身のリン酸化であり、それ以外はEGFRと同様の物質のリン酸化が推測された。 K664ではEGFによるリン酸化は誘導されなかった。 3キメラレセプターとGAPの結合 EGF刺激前後の細胞株3T3sub、EGF1およびEGP1のtotal cell lysateを抗EGFRモノクローナル抗体で免疫沈降し、抗GAPポリクローナル抗体でウエスタンブロット法を行い、EGF1およびEGP1でEGF刺激依存性に120kDaの大きさのバンドを認めた。また抗P-Tyrモノクローナル抗体でウエスタンブロットし、EGF刺激後のEGF1で170kDaに、EGP1で150kDaに著名なリン酸化の亢進を認めた。これはともにチロシンリン酸化されたEGFRおよびEGPR自身と考えられた。さらにEGF刺激後のEGF1およびEGP1の両者で120kDaのリン酸化亢進を認め、EGFRまたはEGPRに結合したGAPと推測された。同じサンプルを抗GAPポリクローナル抗体で免疫沈降し、抗GAPポリクローナル抗体でウエスタンブロット法を行い、GAPの発現量が細胞株間で差がないこと、および上述の120kDaのバンドとGAPのバンドの大きさが一致することを確認した。 また同サンプルで抗GAPポリクローナル抗体で免疫沈降し、抗P-Tyrモノクローナル抗体でブロットして、EGF刺激後にEGF1およびEGP1で62、120、190kDaのチロシンリン酸化バンドを確認した。それぞれp62、GAP、p190と考えられる。GAPの発現および大きさは同サンプルを抗GAPポリクローナル抗体で免疫沈降した後、抗GAPポリクローナル抗体でウエスタンブロット法を行い、確認した。100kDaのバンドは最近クローニングされたp100GAPと考えられる。 上記と同様の方法によりEGPRとPhospatidilinositol 3-Kinase、およびAsh/Grb2との関係を調べたが、EGPRとの結合は認められなかった。 以上の結果よりEGF刺激により、EGFRおよびEGPRがチロシンリン酸化を受けたGAPとそれぞれ結合することが確認された。 4細胞増殖曲線 次に低FCS濃度で10ng/mlのEGF存在下でEGF1、EGP1、およびK664を培養すると、6日目で3T3subに比べてEGF1で約4倍、EGP1で約3倍の細胞数の増加が観察された。これより、EGPRはリガンドの存在下で自己リン酸化を受け、シグナルの少なくとも一部はGAPを介して伝えられ、細胞増殖に関与すると推測された。 ここまでのEGPRの解析より、Ephは未知のリガンド刺激によりシグナルの一部はGAPを介し下流へ伝達され、また細胞増殖に関与することが示唆された。 5Ephトランスジェニックマウスの作成 次にCytomegalovirus(CMV)のエンハンサーおよびchicken 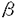 -actinのプロモーターを調節領域として持つmammarian expression vectorであるpUC-CAGGSにEph cDNAを組み込み、CMVのエンハンサー、chicken -actin、Eph cDNA、rabbit -globlinを含む約5kbのfragmentをマウスBDF1受精卵雄性前核にマイクロインジェクションし、Ephトランスジェニックマウスを作成した。得られた5匹のファウンダーマウスから子孫に安定してトランスジーンを伝える2ラインを確立し、実験に用いた。 -actinのプロモーターを調節領域として持つmammarian expression vectorであるpUC-CAGGSにEph cDNAを組み込み、CMVのエンハンサー、chicken -actin、Eph cDNA、rabbit -globlinを含む約5kbのfragmentをマウスBDF1受精卵雄性前核にマイクロインジェクションし、Ephトランスジェニックマウスを作成した。得られた5匹のファウンダーマウスから子孫に安定してトランスジーンを伝える2ラインを確立し、実験に用いた。 6Ephトランスジェニックマウスの解析 Ephトランスジェニックマウスより臓器を摘出し、抗C-Ephポリクローナル抗体によるウエスタンブロット法で脳と心筋でEphの発現を確認した。また同組織でEphのチロシンリン酸化の上昇も認めた。さらに脳および心筋でEphとGAPとの結合を確認した。しかしこれまでの観察では同組織で病理学的には異常所見は認められていない。 以上の結果よりEphとGAPの生体内での結合およびチロシンリン酸化が確認され、細胞株EGP1の結果と矛盾しない所見を得た。 7考案 以上実験より次のような結果を得た。 1)EGFレセプター(EGFR)の細胞外領域cDNAとEphの細胞内領域cDNAを結合したキメラレセプター(EGPR)cDNAを作成し、内因性EGFRを発現していない細胞株NIH3T3sublineにトランスフェクトし、EGPRを高発現する変異安定株EGP1を作成した。 2)EPhのATP binding site(Lys664→Met)を変異させたEphK664を作成し、さらにEGFRとEPhK664をキメラ結合させたEGPRK664を構築した。またEGPRK664を安定発現する細胞株K664を作成した。 3)EGP1はEGF刺激により、EGPR自身および細胞内タンパクのチロシンリン酸化を亢進する。K664はEGF刺激によるチロシンリン酸化の変化を受けない。 4)EGPRはEGF刺激依存性にGTPase activating protein(GAP)と結合し、またGMP自身はチロシンリン酸化を受けた。Phosphatidilinositol 3-Kinase(PI3K)、およびAsh/Grb2とEGPRの結合は確認できなかった。 5)EGP1は親株NIH3T3sublineに比べて低BS濃度培養下でEGF刺激により、6日目で約3倍の細胞増殖を認めた。 6)CMVのエンハンサー、-actinのプロモーターを調節領域として持つEphトランスジェニックマウスにおいて、心筋、脳でEphの発現およびチロシンリン酸化を確認した。しかし組織学的には変化は認められなかった。 7)Ephトランスジェニックマウスの心筋と脳において、EphとGAPの結合を確認した。 Ephの細胞内情報伝達の系路、すなわちEGPRのそれと置き換えることができるが、今回の実験で、EGPRとGAPの結合、すなわちEphとGAPの結合が確認され、これまでの報告とあわせ考えて、何らかのEphリガンド刺激→Eph→GAP→ras→(raf→)核へシグナル伝達の流れがあると推定される。細胞増殖にrasの活性化が関与することは一般に知られており、EphとGAPの結合があることよりEphのシグナルは細胞増殖に関達している可能性が考えられた。 EGF刺激によりEGPRと3T3subの間に細胞増殖の差ができることから、EGPRのシグナル、すなわちEphのシグナルが細胞増殖に関与することが示唆された。EGF刺激依存性にEGPRがGAPと結合し、チロシンリン酸化が上昇する知見と合わせ考え、少なくとも刺激の一部がGAPを介する系で下流へ伝達され、細胞増殖に関わるシグナルに関与していると推測された。これまで明らかでなかったEphの生物学的活性に新しい知見を与えると思われる。 8今後の展望 Ephファミリーは受容体型チロシンキナーゼの中で、細胞内情報伝達機構や生物学的活性がほとんど究明されていないキナーゼの一つである。しかしEphファミリーの発現が主に脳神経経に組織分布していること、および今回のEphの細胞増殖への関与の知見から、Ephファミリーが神経系の発生、分化の制御に関与していることが推測され、その役割はますます興味深いものに成りつつある。 さらに最近Ephファミリーのリガンドが単離され、今後急速に機能解析が発展することが期待される。 またEphトランスジェニックマウスにおいては、発現の時期や組織特異性を検討できるプロモーターの調節を加えることやEphファミリーリガンドの刺激を加えるなどして、Ephの生物学的活性をさらなる究明が可能と考える。 |