血糖値の恒常性を保つための重要なホルモンであるインスリンは、血糖値の上昇に伴って膵ランゲルハンス島の 細胞から分泌される。グルコースは、細胞のインスリン分泌応答において、生理的に最も重要だと考えられる分泌刺激物質であるが、グルコースに応答してインスリン分泌が起こるメカニズムとしては、現在のところ次のような仮説が有力視されている。 細胞から分泌される。グルコースは、細胞のインスリン分泌応答において、生理的に最も重要だと考えられる分泌刺激物質であるが、グルコースに応答してインスリン分泌が起こるメカニズムとしては、現在のところ次のような仮説が有力視されている。 すなわち、グルコーストランスポーターを介してグルコースが細胞内に取り込まれ、まず解糖系やTCAサイクルにより代謝を受けると考えられている。次に、代謝により生成したATPが細胞膜に存在するATP感受性K+チャンネルに作用し、K+チャンネルを閉鎖することが想定されている。このATP感受性K+チャンネルの閉鎖により細胞膜が脱分極し、次に電位依存性Ca2+チャンネル(VDCC)が活性化されると考えられている。そして、Ca2+チャンネルの活性化の結果、細胞内にCa2+が流入し細胞内Ca2+濃度の上昇が起こり、インスリン分泌につながるというものである。これは、1984年のCookらとAshcroftらによる膵細胞におけるATP感受性K+チャンネルの発見に基づくものであり、現在のところ最も有力視されている仮説である。 一方、1993年に岡本らにより提出された仮説では、これとは全く異なる新たな情報伝達経路が想定されている。彼らによると、グルコースに応答してNAD代謝物の一種であるcyclic ADP-ribose(cADP-ribose)という物質が生成し、小胞体からのCa2+の放出を引き起こす可能性が示されている。すなわち、この仮説では、細胞外からのCa2+の流入ではなく、細胞内からのCa2+の動員がインスリン分泌の引き金になることが提案されており、実際にどちらの仮説に沿った経路が働いているのかは判然としていないのが現状である。 ところで、Nicotinamideのアナログである6-Aminonicotinamide(6AN)という物質が、ラットに高血糖を引き起こすという報告がある。これは、ATP感受性K+チャンネル説が提出される以前の報告で、血糖値上昇のメカニズムに関しては明確な考察はなされていない。しかし、NicotinamideがNADの前駆体であることから、この物質がNAD代謝に影響を与える可能性が予想され、細胞のインスリン分泌機構を知る上で利用できる可能性があると考えられる。そこで、この物質の作用点の解明を目標として以下の検討を行なった。 1.6ANによる血糖値の上昇とインスリン分泌応答の低下 6ANをラットに腹腔内投与し、血糖値の変化を測定したところ、投与6-9時間後をピークとする血糖値の上昇が観察された。そこで、この血糖値の上昇作用が、膵臓からのインスリン分泌反応の低下によるものであるかどうかを調べるために、6AN処理ラットにグルコースを経口負荷し、グルコース負荷時にインスリン分泌応答の低下が観察されるかどうかを調べた。その結果、6AN処理ラットでは、グルコース負荷によるインスリン分泌応答に低下が観察され、膵細胞からのインスリン分泌反応が障害を受けている可能性が示唆された。 そこで、次に6AN処理ラットからランゲルハンス島を単離しin vitroでグルコースを作用させてインスリン分泌応答を測定した結果、やはり細胞自体のインスリン分泌応答に抑制が見られることが明らかになった。 in vivoで見られた6ANの作用は、in vitroで作用させた場合にも観察された。すなわち、ランゲルハンス島あるいはインスリン分泌応答を持つ培養細胞株であるMIN6細胞を6ANで前処理することにより、グルコース刺激に伴うインスリン分泌応答が抑制されることが明らかになった。 2.細胞内Ca2+濃度変化とインスリン分泌応答-6AN処理細胞におけるCa2+依存性応答の消失- グルコース刺激によるインスリン分泌応答においては、細胞内Ca2+濃度([Ca2+]i)の上昇が重要な役割を担っていると考えられている。そこで、MIN6細胞をグルコースで刺激し、経時的な[Ca2+]iの変化を測定した。その結果、6AN処理細胞では、グルコース刺激に伴う[Ca2+]iの上昇が観察されないことが明らかになった。このことから、6AN処理細胞では、[Ca2+]iの上昇以前の段階に障害が起きており、そのためにインスリン分泌応答が抑制されている可能性が示唆された。 一方、無処理細胞を用いて、グルコース刺激に伴う[Ca2+]iの変化の性質を詳しく解析したところ、[Ca2+]iの上昇までには約2分のlag timeがあること、細胞外Ca2+の非存在下では[Ca2+]iの上昇が観察されないこと、VDCCの阻害剤により[Ca2+]iの上昇が消失することが明らかになった。また、ヨード酢酸などの解糖系の阻害剤を用いた結果から、[Ca2+]iの上昇がグルコース代謝に依存したものであることが明らかになった。 これらの結果から、グルコース刺激に伴う[Ca2+]iの上昇反応は、グルコース代謝、ATP感受性K+チャンネルの閉鎖、脱分極、VDCCの活性化、Ca2+の流入という仮説に沿った反応である可能性が示唆された。 一方のcADP-riboseによるCa2+の動員の可能性に関しては、グルコース刺激によりcADP-ribose濃度の上昇が観察されないこと、cADP-riboseの合成酵素の強制発現により[Ca2+]iに変化が観察されないことなどの事実が明らかになり、[Ca2+]iの上昇応答に関与している可能性は少ないことが明らかになった。 次に、グルコース刺激時に[Ca2+]iの変化とインスリン分泌応答を同時測定することにより、両者の関係を対応させることを試みた。その結果、新たな事実が明らかになった。グルコース刺激後、[Ca2+]iの上昇までには約2分のlag timeが観察されたが、インスリン分泌応答にはlag timeが存在せず、[Ca2+]iの上昇に先行する一過性のインスリン分泌反応が起こることがわかった。このインスリン分泌反応は、細胞外Ca2+の非存在下でも観察され、[Ca2+]iの上昇に先行する反応であることから、Ca2+非依存性のインスリン分泌応答であることが明らかになった。 一方、この一過性の分泌反応の後に、約5分後から緩やかで持続的なインスリン分泌反応が起こることがわかった。この持続的なインスリン分泌反応は、グルコースで刺激している間継続的に観察され、インスリン分泌量の総量としては前のものをはるかに上回っていた。こちらに関しては、細胞外Ca2+の非存在下では観察されないこと、[Ca2+]iの上昇が起こらない条件下では観察されないこと、代謝依存性があることなどの性質が、従来の仮説と一致しており、グルコース代謝に基づく、Ca2+の流入を介するインスリン分泌応答であることが示唆された。 ところで、6AN処理細胞では、最初の一過性のインスリン分泌応答は影響を受けずに、次のCa2+の流入を介するインスリン分泌応答だけが抑制されていることが明らかになった。このことから、6AN処理細胞では、グルコース代謝に基づく、Ca2+依存性のインスリン分泌応答だけが障害を受けていることが明らかになった。 そこで、6AN処理細胞において、グルコース刺激からCa2+の流入に至る経路のどこに障害があるのかを検討した。この目的のために、ATP感受性K+チャンネルに直接作用し、脱分極とVDCCの活性化を介してCa2+の流入を引き起こすGlibenclamideを用いた。その結果、Glibenclamideで刺激した場合には、6AN処理細胞においても、[Ca2+]iの上昇とインスリン分泌応答が観察されることがわかった。このことから、6AN処理細胞のATP感受性K+チャンネルからインスリン分泌応答に至る経路は正常に機能していることが明らかになった。したがって、6AN処理により障害を受けるのはATP感受性K+チャンネル以前のグルコース代謝の段階である可能性が示唆された。 3.NAD量の減少によるグルコース代謝の抑制 ところで、6AN処理細胞の細胞内ヌクレオチドを分析してみると、正常なNADが減少し、代わって6ANのNADアナログ(N’AD)が出現することが明らかになった。インスリン分泌応答に関与することが示されたグルコース代謝の段階には、解糖系と、TCAサイクルにNAD依存性の酸化反応があることから、6AN処理によりこの段階が障害を受け、インスリン分泌応答が抑制されている可能性が考えられた。 そこで、グルコースから産生されるCO2量を測定することにより、グルコースの代謝活性を測定したところ、6AN処理細胞では、無処理細胞の約5分の1にまで活性が低下していることが明らかになった。 また、無処理細胞では、グルコース刺激による細胞内NAD(P)H濃度の上昇が観察されたが、6AN処理細胞ではこの変化が全く起こらないことが明らかになった。これらの結果から、6AN処理細胞では、グルコース代謝におけるNAD依存性の酸化反応が進行せず、そのためにグルコース代謝が抑制されている可能性が明らかになった。 ところで、6AN処理細胞ではNAD量の減少とN’ADの生成という二つの現象が起きていることから、どちらが6ANの持つ抑制作用の原因であるのかを検討した。この目的のために、6ANを一度作用させた後、洗浄除去した細胞を利用した。6AN洗浄除去細胞では、NAD量は回復するが、N’ADが残存することから、N’ADのみの効果を調べることができた。この細胞をグルコースで刺激し、[Ca2+]iの変化を観察したところ、6AN処理細胞の場合とは異なり、グルコース刺激に応答して[Ca2+]iの上昇が起こることがわかった。このことから、グルコース刺激による[Ca2+]iの上昇応答に対して、N’ADは積極的な阻害作用を持たないことがわかった。したがって、6AN処理細胞で観察されたグルコースに対する応答の消失は、NAD量の減少によるものであることが明らかになった。 以上の結果を下図にまとめた。6AN処理細胞では、グルコース刺激に応答したCa2+の流入を介するインスリン分泌応答に低下が観察された。しかし、K+チャンネルに直接作用するGlibenclamideに対する応答は保持されていたことから、K+チャンネル以前のグルコース代謝の段階が傷害を受けている可能性が考えられた。そこでグルコース代謝について検討したところ、6AN処理細胞ではグルコース代謝におけるNAD依存性の反応が抑制されている可能性が明らかになった。このような代謝抑制作用は、6ANにより細胞内NAD量が減少するために起こることがわかった。 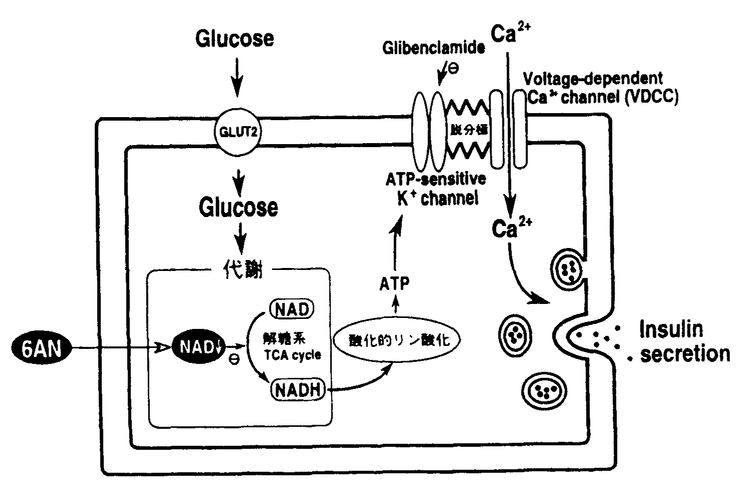 図表 図表 |