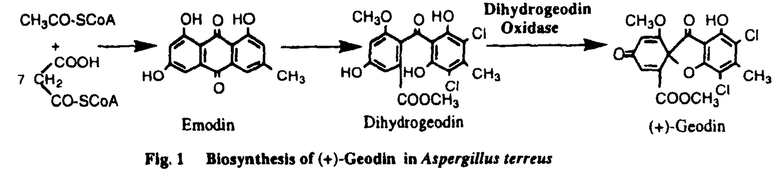
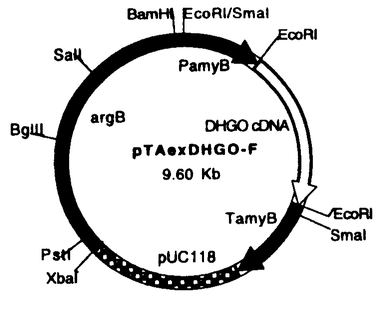
フェノール酸化的縮合反応はmorphineなどの生合成に見られるように二次代謝産物の骨格形成に関わる重要な生合成反応の一つである。我々は、これまで菌類二次代謝産物の生合成を酵素、遺伝子レベルで解明することを目的に検討を行ってきたが、Aspergillus terreusの生産する抗真菌活性物質(+)-geodin生合成の最終段階は立体特異的分子内フェノール酸化的縮合反応であり、当研究室において、この反応を触媒する酵素dihydrogeodin oxidase(DHGO)が精製され、ブルー銅タンパクであることが明らかにされている。(Fig.1)私は、本研究においてdihydrogeodin oxidaseの一次構造、触媒するフェノール酸化的縮合反応の反応機構、および、二次代謝酵素系の発現制御などの解析を目的に本酵素遺伝子のクローニングを試みた。  Fig.1 Biosynthesis of(+)-Geodin in Aspergillus terreus(1)Dihydrogeodin oxidase遺伝子のクローニング Fig.1 Biosynthesis of(+)-Geodin in Aspergillus terreus(1)Dihydrogeodin oxidase遺伝子のクローニング 抗DHGOポリクローナル抗体を用いて、A.terreus cDNAライブラリーをスクリーニングし、4個の陽性クローンを選択した。各インサートの塩基配列を決定したところ、いずれもコード領域は短かったものの酸化酵素関連の同一タンパクをコードしているものと考えられた。そこで、このcDNAをプローブとしてゲノミックサザン分析により検討したところ、5.5kb SacI断片中にDHGO遺伝子全長が存在していると推定され、このゲノムDNA断片をクローニングした。得られたクローンについてDHGOをコードしていると推定される領域の塩基配列を決定し、全長2.5kbにわたるopen reading frameの存在を確認した。(Fig.2)この遺伝子は、糸状菌のラッカゼ、植物のアスコルビン酸酸化酵素遺伝子との間にアミノ酸レベルで36%と29%の相同性が見られたこと、また、銅タンパクに見られる銅結合モチーフが存在していることから、ブルー銅タンパクであるDHGOをコードしているものと考えた。また、open reading frame中に複数のイントロンの存在が推定されたことから、改めてcDNAのクローニングを行った。  Fig.2.Restriction map of 5.5 kb DNA SacI fragment(A)and cDNA clons(B)from Aspergillus terreus.cDNA1,2,3 4 were cloned by cDNA library screening; cDNA 5 was cloned by PCR,White regions indicate introns. Black boxes indicate exons. Fig.2.Restriction map of 5.5 kb DNA SacI fragment(A)and cDNA clons(B)from Aspergillus terreus.cDNA1,2,3 4 were cloned by cDNA library screening; cDNA 5 was cloned by PCR,White regions indicate introns. Black boxes indicate exons.  ZAPIIベクターを用いてA.terreusより、新しくcDNAライブラリーを構築し、先にクローニングした5.5kb SacI断片の内、DHGOをコードしていると思われる2.5kb BamHI断片をプローブとして、約4万個のクローンをスクリーニングし、4個の陽性クローンを得た。この中で最長のインサートは1.6kbであったが、精製タンパクの分子量およびノーザン分析の結果より、これはDHGOcDNAの全長ではないと考えられた。そこで、DHGOゲノム遺伝子の5’側配列を利用してPCR法により全長2.5kbのDHGOcDNAをクロニーングし、全塩基配列を決定した。その塩基配列より推定したアミノ酸配列をFig.3に示す。クローニングされたcDNAとゲノムDNAの塩基配列の比較から、6つのイントロンの存在が確認された。いずれも5’-GT,3’-AGのconsensus配列が認められた。また、5’側上流にはCAAT box,TATA boxと考えられる配列も存在している。今回クローニングした遺伝子は、アミノ酸605個からなる分子量68KDaのタンパク質をコードしており、精製したDHGOのサブユニットの分子量76KDaとは差があるものの、これは翻訳後の糖鎖の付加によるものと考えられる。また、精製DHGOのN末アミノ酸配列が認められたこと、銅結合モチーフが存在していることを合わせ考えると、本遺伝子がDHGOをコードするものと判断した。これを確認するために、異種糸状菌での発現を試みた。 ZAPIIベクターを用いてA.terreusより、新しくcDNAライブラリーを構築し、先にクローニングした5.5kb SacI断片の内、DHGOをコードしていると思われる2.5kb BamHI断片をプローブとして、約4万個のクローンをスクリーニングし、4個の陽性クローンを得た。この中で最長のインサートは1.6kbであったが、精製タンパクの分子量およびノーザン分析の結果より、これはDHGOcDNAの全長ではないと考えられた。そこで、DHGOゲノム遺伝子の5’側配列を利用してPCR法により全長2.5kbのDHGOcDNAをクロニーングし、全塩基配列を決定した。その塩基配列より推定したアミノ酸配列をFig.3に示す。クローニングされたcDNAとゲノムDNAの塩基配列の比較から、6つのイントロンの存在が確認された。いずれも5’-GT,3’-AGのconsensus配列が認められた。また、5’側上流にはCAAT box,TATA boxと考えられる配列も存在している。今回クローニングした遺伝子は、アミノ酸605個からなる分子量68KDaのタンパク質をコードしており、精製したDHGOのサブユニットの分子量76KDaとは差があるものの、これは翻訳後の糖鎖の付加によるものと考えられる。また、精製DHGOのN末アミノ酸配列が認められたこと、銅結合モチーフが存在していることを合わせ考えると、本遺伝子がDHGOをコードするものと判断した。これを確認するために、異種糸状菌での発現を試みた。  Fig.3 The deduced amino acid sequence of dihydrogeodin oxidase from Aspergillus terrues.Copper binding moltifs are boxed;N-terminal amino acid sequence of dihydrogeodin oxidase is underlined.(2)Dihydrogeodin oxidaseの異種糸状菌での発現 Fig.3 The deduced amino acid sequence of dihydrogeodin oxidase from Aspergillus terrues.Copper binding moltifs are boxed;N-terminal amino acid sequence of dihydrogeodin oxidase is underlined.(2)Dihydrogeodin oxidaseの異種糸状菌での発現 5.5 kb SacI断片中には、DHGO遺伝子自身のプロモーターの存在が考えられ、この遺伝子断片を直接異種糸状菌に導入し、発現を試みた。まず、遺伝子工学系が確立しているAspergillus nidulansを用いたところ、全く発現は認められなかった。その原因は、二次代謝系の遺伝子である本遺伝子のプロモーターがA.nidulans中で機能しなかったものと考え、次に、A.terreusと類似の二次代謝系を有するPenicillium frequentansをホストとして用いることとした。5.5kb SacI断片を組み込んだプラスミドpDHGOをhygromycin B耐性遺伝子を持つpDH25と共にP.frequentansにco-transformationし、得られたhygromycin B耐性株についてサザン分析を行い、DHGO遺伝子がP.frequentansのゲノムに組み込まれた形質転換体を選択した。この形質転換体についてWestern blottingで分析したところ、A.terreusのDHGOと同一分子量をもつタンパクとして発現されていることを確認することができた。(Fig.4)その活性については、P.frequentansのsulochrin oxidaseとの分離が難しく、確認は困難であった。そこで、DHGO genomic DNA、cDNAを糸状菌の発現ベクターpTAexのTaka amylase Aのプロモーター下流に組み込み(Fig.5)、A.nidulans argB mutantに導入した。得られた形質転換体を4日間培養後、粗酵素液を調製して、DHGO酵素活性を測定した。逆向きにgenomeDNA、cDNAを組み込んだ形質転換体では、全く活性は見られなかったのに対し、正しい向きにcDNAを組み込んだ形質転換体において、DHGO酵素活性を確認することができた。この酵素反応の生成物が(+)-geodinであることはHPLC、MS等で確認した。  図表Fig.4 Western blot analysis of P.frequentans pDH25 and pDHGO co-transformants A:P.frequentans; B: Aspergillus terreus T1-T5: P.frequentans pDH25 and pDHGO co-transformants / Fig.5 Experssion vector for DHGO(3)Aspergillus terreus とPenicillium frequentansの形質転換系の確立 図表Fig.4 Western blot analysis of P.frequentans pDH25 and pDHGO co-transformants A:P.frequentans; B: Aspergillus terreus T1-T5: P.frequentans pDH25 and pDHGO co-transformants / Fig.5 Experssion vector for DHGO(3)Aspergillus terreus とPenicillium frequentansの形質転換系の確立 近年、Aspergillus nidulansを中心に糸状菌の遺伝子工学系の開発が進められ、糸状菌における選択マーカーとしては薬剤耐性遺伝子や栄養要求性マーカーなどが知られている。P.frequentansは200 g/mlの濃度でhygromycin B感受性であり、hygromycin B耐性遺伝子を持つプラスミドpDH25、またpAN7-1を持ちい、P.frequentansのプロトプラストの形質転換系を確立した。その形質転換効率は1gDNAあたり、10個から20個の形質転換体であった。Aspergillus terreusにおいてはA.oryzae niaD遺伝子を持つプラスミドpSTA14をAspergilus terreus niaD mutantに導入し、1-5transformants/g DNAの形質転換系を確立した。 g/mlの濃度でhygromycin B感受性であり、hygromycin B耐性遺伝子を持つプラスミドpDH25、またpAN7-1を持ちい、P.frequentansのプロトプラストの形質転換系を確立した。その形質転換効率は1gDNAあたり、10個から20個の形質転換体であった。Aspergillus terreusにおいてはA.oryzae niaD遺伝子を持つプラスミドpSTA14をAspergilus terreus niaD mutantに導入し、1-5transformants/g DNAの形質転換系を確立した。 (4)まとめ 私は本研究において立体特異的分子内C-Oフェノール酸化的縮合反応を触媒する酵素dihydrogeodin oxidaseのゲノムDNAおよびcDNAをクローニングし、活性を有する酵素タンパクとしてA.nidulansで発現させることに成功した。植物のmorphine、 berbamunine生合成における分子内C-CおよびC-Oフェノール酸化的縮合反応については、酵素、遺伝子レベルで解析が進められており、これがP-450により、触媒されることが報告されている。しかしながら、菌類の二次代謝産物生合成における立体特異的分子内C-Oフェノール酸化的縮合反応については、これを触媒する酵素の遺伝子クローニングに成功したのは本研究が初めてである。機能的には類似の反応が、植物においては、P-450により触媒され、一方、糸状菌においては銅タンパクによることは興味深い。また、ブルー銅タンパクとして、本酵素タンパク中における銅の存在状態と、その触媒する反応機構にも興味が持たれるが、本酵素の発現系が確立できたことは、今後、その解明に大きく寄与できるものと期待される。また、DHGO遺伝子のプロモーターは、A.nidulansでは機能せず、A.terreusと類似の二次代謝系を持つP.frequentansで機能することから、本遺伝子のプロモーターの解析は、糸状菌における二次代謝発現の制御機構を探る手がかりとなること、また、アフラトキシンやメラニンなど、糸状菌の二次代謝産物生合成遺伝子クラスターの存在が予想されていることから、今回クローニングしたDHGO遺伝子近傍に存在すると考えられる(+)-geodin生合成に関わるポリケタイド合成酵素、酸化酵素、メチルトランスフェラーゼなどの遺伝子を同定し、生合成系全体を遺伝子レベルで解明することが期待される。 |