肝臓は、血糖の恒常性を維持していく上で非常に重要な臓器である。肝臓のグリコーゲン代謝は様々な神経やホルモンの支配によって複雑に制御されている。 1982年、岡島、宇井は、ラット肝細胞を初代培養するとNorepinephrine刺激によるGlycogen Phosphorylase(Phosphorylase)活性化の受容体依存性が 1受容体から 1受容体から 2受容体へと機能転換することを見出し、これを「→転換」と称して報告した。この現象は動物の生体内においても、肝臓の外科的、化学的な損傷時に発現することが知られている。 2受容体へと機能転換することを見出し、これを「→転換」と称して報告した。この現象は動物の生体内においても、肝臓の外科的、化学的な損傷時に発現することが知られている。 また、梶山らは、この「→転換」が、分離した肝細胞の初代培養において低細胞密度培養時にのみ発現してくることを見出した。また、この培養時に肝細胞の膜画分を添加することによって「→転換」の発現が抑制されるという知見を得た。さらに、膜画分の添加によるこの現象の発現抑制と、肝細胞のDNA合成の抑制に相関性があることから、「→転換」は細胞増殖期への移行(G0/G1→S)に伴う現象であることを示した。 本研究では、肝細胞の「→転換」に伴って起こる膜レベル以降の細胞内情報伝達系の変化、特に細胞内Ca2+動員系の変化について解析を行なった。 まず、肝細胞のホルモン刺激における最終応答と考えられるPhosphorylaseの活性化について検討を行なった。Phosphorylaseは、肝臓のグリコーゲン分解によるグルコース産生に非常に重要な酵素であり、Phosphorylase Kinaseによるリン酸化によって活性化されることが知られている。この酵素活性はcAMPとCa2+の両方で制御されており、これら二つのセカンドメッセンジャーの細胞内濃度上昇により活性化される。ラット遊離肝細胞を高細胞密度(1.6×105cells/cm2)と低細胞密度(2×104cells/cm2)で初代培養し、各培養時間において受容体刺激によるIP3生成とPhosphorylaseの活性化について調べた。高細胞密度培養においては、培養時間の経過と共に受容体刺激によるIP3生成が変化することはなかった。これに対し、低細胞密度培養時には培養時間の経過に伴って比較的ゆっくりと減少していくことが観察された。また、PhosphorylaseのCa2+動員性のホルモン刺激による活性化は、「→転換」に伴い低細胞密度培養時にのみ急速に減少していくことが観察された。この受容体刺激によるIP3生成とPhosphorylaseの活性化は、同じ受容体刺激を行なっているにも関わらず、培養による減少の時間経過が異なっていた。つまり、「→転換」に伴う受容体刺激によるIP3生成の減少がほとんど観察されない培養時間において、同じ刺激によるPhosphorylaseの活性化は遊離肝細胞の70%近くまで減少していることが示された。 このIP3生成とPhosphorylase活性化の時間的な相違はどのレベルで起こっているかということについて、以降検討を行なった。前にも述べたように、Phosphorylaseの活性化を行なうPhosphorylase KinaseはcAMPとCa2+の細胞内濃度上昇によって活性化されることが知られているので、細胞内からのCa2+動員系が「→転換」に伴って変化していることが想定された。また、細胞を破砕した実験の結果から、Phosphorylaseはホルモン刺激により細胞内ストアから動員されたCa2+によって大部分が活性化されることが示唆された。そこで、細胞外液にCa2+を含まない状態で受容体刺激を行ない、細胞内ストアからのCa2+動員を各培養時間経過した細胞で観察した。高細胞密度培養では、培養時間の経過に伴って受容体刺激による細胞内からのCa2+動員が変化することはなかった。これに対し、低細胞密度培養時には、培養時間の経過に伴い受容体刺激による細胞内からのCa2+動員は急速に減少していくことが観察された。この受容体刺激による細胞内からのCa2+動員の「→転換」に伴う減少は、同じ刺激によるPhosphorylase活性化減少の時間経過と一致していることが示された。 このことから、「→転換」に伴い、小胞体中のIP3感受性Ca2+ストアに何らかの変化が起こっている可能性が考えられた。そこで、肝細胞の小胞体を含む画分においても、同様の現象が観察されるかどうか検討を行なった。遊離肝細胞を低密度と高密度で初代培養した後ホモジェネート(20000×g遠心操作後の上清)を調製し、この画分に含まれる小胞体のCa2+ストアからのIP3感受性Ca2+放出の測定を膜透過性のないFura-2を用いて行なった。高密度培養の細胞から調製した画分からのIP3添加によるCa2+放出は培養時間の経過に伴って変化することはなかったが、低密度培養の細胞から調製した画分でのCa2+放出では、IP3の濃度依存性が高濃度側に移行しており、またIP3の添加によるCa2+放出能も低下していることが観察された。さらに、この画分に対するIP3の結合活性を測定したところ、高細胞密度で培養を行なった場合にはIP3に対する親和性や結合部位数が変化することはなかったが、低細胞密度で培養を行なった場合にのみ、培養時間の経過に伴って急速に結合部位数が減少していくことが示された。この結果から、「→転換」の初期に観察される受容体刺激によるIP3生成と細胞内からのCa2+動員の時間的なずれは、IP3結合部位の減少を伴う、IP3感受性Ca2+プールからのIP3に応答したCa2+放出の減少に起因することが示された。 様々な細胞の機能に重要な役割を果たす細胞内Ca2+濃度上昇は、細胞内からの動員ばかりではなく、細胞外からの流入によっても引き起こされる。そこで、受容体刺激による細胞外からのCa2+流入を、「Ca2+の後添加法」を用いることにより細胞内からの動員と分離して測定し、細胞外からのCa2+流入が「→転換」に伴ってどのように変化するかについても検討を行なった。高細胞密度培養時には細胞外からのCa2+流入が培養時間の経過と共に変化することはなかった。これに対して、低細胞密度培養時には、細胞外からのCa2+流入が培養時間の経過と共に減少していくことが観察された。しかし、この減少は、Phosphorylase活性化の「→転換」に伴う減少の時間経過と異なり、むしろ受容体刺激によるIP3生成の減少の時間経過と類似していた。また、受容体作動性のCa2+流入(Receptor-operated Ca2+influx)とは独立したCa2+流入経路として想定されている貯蔵部位直結性のCa2+流入(CapacitativeCa2+influx)は肝細胞ではほとんど観察されず、また「→転換」に伴う変化も観察されなかった。このことから、細胞外からのCa2+流入の「→転換」に伴う減少は、膜上の受容体数の「→転換」に伴う減少と相関していることが考えられた。また初代培養肝細胞においては、細胞外からのCa2+流入は細胞内のCa2+プールが枯渇したことによって起こる二次的なものではなく、細胞内からのCa2+動員とは独立したCa2+動員経路である可能性が考えられた。 また、新たに肝細胞の小胞体を含む画分において、Cyclic ADP-ribose(cADPr)の添加によりCa2+放出が起こることを見出した。このCa2+放出はcADPrに特異的であり、他の類似した化合物ではCa2+放出が起こらなかった。またHeparinの添加効果から、cADPrはIP3感受性のCa2+ストアとは独立した別のストアからCa2+を放出させることが示された。ただし、このストアはRyanodine感受性のストアではない可能性が考えられた。また、cADPrの添加によるこのストアからのCa2+放出は、肝細胞の「→転換」に伴って変化することはなかった。さらに、cADPrがホルモン刺激によって生理的にCa2+放出を起こすメッセンジャーである可能性を考え、様々なホルモン刺激による細胞内のcADPr濃度変化について検討を行なったが、刺激による有意な濃度上昇は観察されなかった。 これらのことから、肝細胞の「→転換」に伴って、ホルモン刺激によるIP3生成の培養時間に伴う減少に先行して、同じ刺激による細胞内ストアからのCa2+動員が減少していることが示された。これは、小胞体上のIP3結合部位の減少を伴った、IP3感受性Ca2+ストアからのCa2+放出が減少することに起因すると結論づけられた。また、肝細胞にcADPr感受性のCa2+ストアが存在することが示された。 | 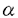 →
→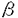 転換」に伴う細胞内カルシウムの変化
転換」に伴う細胞内カルシウムの変化