非相同的組換えはDNAのほとんど相同性のない部位の間で起こる遺伝的組換えであり、DNAの挿入、重複、欠失、転座などをひきおこすことが知られている。このDNAの再編成は、生物の進化にも関係するが、その一方で、癌化した細胞で見られる染色体異常の原因の一つとしても考えられるなど、生物の生存に対して潜在的な危険性もはらんでいる。また、動物細胞においては、導入したDNAが非相同的組換えによりランダムに組み込まれてしまい、遺伝子ターゲティングを困難にしている。 真核生物の非相同的組換えの機構を明らかにするため、分裂酵母における遺伝子ターゲティングのモデル系を作製し、導入したDNAが染色体に組み込まれた非相同的組換え体を効率よく検出することを可能にした。得られた非相同的組換え体についてその組換え部位の塩基配列を決定したところ、染色体の組換え部位では欠失がほとんど起こらないことを見いだした。この特徴から、非相同的組み込みの特徴として、構造の異なるDNA末端どうしを結合させるEnd Joining蛋白質が働くという新しいモデルを提案した。 このモデルに基づき、DNA修復に関わる突然変異株を用いて、各変異の非相同的組換えへの影響を調べたところ、4つの突然変異株(rad8,rad11,rad21,swi10)で非相同的組換え頻度の相対的な低下が明らかになった。そのうち、rad21突然変異株では非相同的組換え頻度、並びにEnd Joining活性の低下が示され、rad21遺伝子産物はDNA末端どうしを結合させる段階で非相同的組み込みに関与している可能性が示唆された。この結果は、End Joiningモデルを支持するもので、このモデルの確立の基盤となるものである。また、従来からいわれている組換え修復(相同的組換えを介した修復)とは異なる新しいタイプの修復が存在することを示唆するものであり、重要な知見である。 さらにこれまで解析の進んでいなかった分裂酵母のDNA修復遺伝子であるrad6遺伝子の機能を調べるために、rad6遺伝子をクローニングしその塩基配列を決定した。その結果、rad6遺伝子がコードする蛋白質は、出芽酵母の突然変異に関わる修復系に属するRad18蛋白質と相同であることが明らかになった。 本研究では、分裂酵母の非相同的なDNAの組み込みの機構を解析し、End Joiningモデルという新しいモデルを提案し、さらに遺伝学的解析からこのモデルの正しさを裏付ける証拠を得た。さらにこの組み込みに関与する遺伝子rad21に関する結果は、新しいDNA修復系の存在を示唆した。以上の点で分子遺伝学、分子生物学の分野の進展に貢献したと評価される。以上により本論文は博士(薬学)の学位を受けるに充分であると判定された。 | 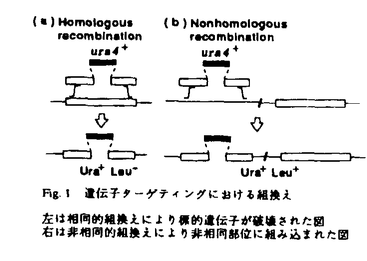


 線に強い感受性を示し、また、DNAの二重鎖切断修復能に欠損があることが知られている。rad21突然変異株での非相同的組換えの相対的な割合の減少が、非相同的組換え頻度の低下と相同的組換え頻度の上昇のいずれによるのかを明らかにするために、相同的組換えに関しては形質転換効率に対する相同的組換え頻度を、非相同的組換えに関しては、染色体と全く相同性をもたないura4+遺伝子断片を導入し、その非相同的組み込みの頻度を形質転換効率で補正することで調べた。その結果、相同的組換えは野生株とほとんど変わらないのに対し、非相同的組換え頻度は野生株に比べて顕著に低下しており、rad21遺伝子産物が非相同的組み込みに関与することが明らかになった(Fig.4)。
線に強い感受性を示し、また、DNAの二重鎖切断修復能に欠損があることが知られている。rad21突然変異株での非相同的組換えの相対的な割合の減少が、非相同的組換え頻度の低下と相同的組換え頻度の上昇のいずれによるのかを明らかにするために、相同的組換えに関しては形質転換効率に対する相同的組換え頻度を、非相同的組換えに関しては、染色体と全く相同性をもたないura4+遺伝子断片を導入し、その非相同的組み込みの頻度を形質転換効率で補正することで調べた。その結果、相同的組換えは野生株とほとんど変わらないのに対し、非相同的組換え頻度は野生株に比べて顕著に低下しており、rad21遺伝子産物が非相同的組み込みに関与することが明らかになった(Fig.4)。
