真核単細胞生物である分裂酵母は、外界の栄養源の枯渇により有性生殖過程に移行する。有性生殖過程においては、P型(h+)とM型(h-)の二種の接合型の細胞が接合して二倍体となり、引き続き減数分裂・胞子形成を行う。接合には接合フェロモンによる細胞間の情報伝達が必要とされ、P型細胞はP-ファクターを分泌し、M-ファクター受容体を産生する。逆にM型細胞はM-ファクターを分泌し、P-ファクター受容体を産生する。接合フェロモンおよびその受容体が発現され、二種の細胞が互いに相互作用する接合過程は、高等生物において細胞が特定の機能を発現し、他の細胞と相互作用する過程と基本的に類似した様相を呈する。申請者は接合フェロモンやその受容体が接合型特異的に発現される機構を細胞分化のモデル系として捉え、その解明を目指して本研究を行った。 分裂酵母の接合型特異的な遺伝子の発現は、細胞の接合型により厳密に決定されると同時に、外界の栄養条件、また相手細胞からの接合フェロモンの受容によっても制御される。細胞の接合型は接合型遺伝子matPcとmatMcによりそれぞれP型、M型に決定されるが、これらの遺伝子産物がどのようにして接合型特異的な遺伝子の発現を制御するのかに関しては従来ほとんど明らかにされてこなかった。接合フェロモンの受容は受容体とその下流のMAPキナーゼ系を経て伝達されるが、この情報が遺伝子の発現に変換される具体的な機構もまた不明のまま残されている。申請者はこれらの点をより明確にするため、M型細胞に特異的に発現されるsxa2遺伝子に着目して、この遺伝子の転写誘導に関与する遺伝子の単離を試みた。sxa2遺伝子はP-ファクターを分解するプロテアーゼをコードし、そのmRNAはM型細胞においてP-ファクターの受容下でのみ観察される。P-ファクター受容の情報伝達に関わるMAPキナーゼをコードするspk1を欠損した細胞では、sxa2遺伝子の発現が見られない。そこで申請者はspk1遺伝子破壊株にsxa2-lacZ融合遺伝子をもたせ、ここに分裂酵母の遺伝子ライブラリーを導入して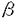 -galactosidase活性の発現が見られるクローンを単離した。 -galactosidase活性の発現が見られるクローンを単離した。 このようにして単離したクローンの一つには398アミノ酸をコードするORFが存在していた。配列から予想されるタンパク質はそのN末端側において、出芽酵母の接合型特異的な遺伝子の転写制御因子であるMCM1、ならびにヒトのc-fosのseruMresponse elementに結合するSeruMResponse Factor(SRF)とそれぞれ36.0%、31.1%の相同性を有した。相同領域はMADS-boxとよばれるモチーフを含んでいた。MADS-boxはDNAへの結合とホモダイマーの形成に必要十分な領域であることが知られており、この遺伝子産物もまた転写因子として機能すると思われた。 申請者は、P型とM型が一つのコロニー内に混在するh90型細胞において、上記の遺伝子を破壊した。遺伝子破壊株は栄養増殖に関してはまったく欠損が見られなかったが、コロニー内で接合・胞子形成できなかった。より詳細に解析した結果、この細胞はP野生型細胞とは接合できるがM野生型細胞とは接合できず、P型特異的な接合不能(maP)の表現型を示すことが明らかとなった。これまでに単離されているmaP変異と比較検討し、この遺伝子は未だクローニングの報告のなかったmap1遺伝子であることを申請者は指し示した。 map1はP型細胞特異的に接合不能を賦与する変異として、分裂酵母の分子遺伝学的研究のごく初期に単離されたものである。map1遺伝子は、その変異株の解析からおそらくP型細胞においてのみ機能し、P-ファクターの産生およびM-ファクター受容体の産生に関与すると考えられてきた。しかし、申請者によってM型特異的に発現するsxa2遺伝子の転写誘導因子としてmap1遺伝子が単離されたことは、従来の概念では説明できないものであった。そこで申請者はM型細胞におけるmap1遺伝子の機能を探るため、map1を欠くM型細胞の表現型を詳細に観察した。その結果M型細胞においてもmap1遺伝子が破壊されると接合能が若干低下することが判明した。次にmap1を欠くM型細胞のP-ファクター存在下でのsxa2の転写誘導をノザン解析により調べた。その結果、map1遺伝子破壊株ではsxa2をはじめとするM型細胞特異的な遺伝子の転写レベルは野生型に比べ低く、map1遺伝子がM型細胞においても有性生殖に必要な遺伝子の発現制御に関与していることを明らかにした。また、map1遺伝子はM型細胞においてもP型細胞と同レベルで発現していたことも、map1遺伝子のM型細胞での機能を裏づけた。 申請者はmap1遺伝子の機能をさらに解析するため、map1を過剰発現するプラスミドを導入した細胞を栄養源飢餓下において、接合型特異的な遺伝子の転写レベルを解析した。P型特異的な遺伝子としてP-ファクターをコードするmap2、M-ファクター受容体をコードするmap3、アグルチニンをコードするmap4の発現を調べた。map1遺伝子破壊株でこれらの遺伝子のmRNAが見られないのに対し、map1を過剰発現したP型細胞においてはM-ファクターの非存在下にもかかわらず高いレベルでの転写が観察された。一方M型特異的な遺伝子としてP-ファクター受容体をコードするmam2、およびsxa2遺伝子の発現を調べた。この場合map1を過剰発現したM型細胞でP-ファクターの非存在下にもかかわらずmRNAの転写が強く誘導されていた。これらの結果からmap1遺伝子が細胞の接合型に則って、接合型特異的な遺伝子の発現を正に制御することを推論した。 申請者は次いで、Map1が接合型遺伝子産物と相互作用する可能性をyeast two-hybrid systeMをもちいて検討した。GAL4 activating domainに融合したMap1とCAL4 DNA-binding domainに融合したMat1-Pcを同時に導入した出芽酵母レポーター株では、GAL4に依存したHIS3遺伝子の転写が認められ、Map1とMat1-Pcの物理的な相互作用が推定された。 以上の結果から、出芽酵母の接合型特異的な遺伝子の発現制御機構と比較しつつ、申請者はMap1の作用機構について次のような一つのモデルを提唱している。出芽酵母においては接合型遺伝子産物の 1あるいは2とMCM1が複合体を形成して、a型および型のいずれの接合型においても細胞特異的な遺伝子の発現を制御している。map1遺伝子産物もまたMCM1と同様に接合型遺伝子産物と相互作用し、P型とM型のそれぞれに特異的な遺伝子の上流配列に結合し、その発現を制御していると申請者は考えている。その際興味深いことに、map1遺伝子はMCM1遺伝子と異なり、細胞分化に特異的に作用し細胞増殖には不要である。またその機能はP型細胞では必須であるが、M型細胞においては絶対に必要なわけでなく、その存在意義は二種の細胞で必ずしも同等ではない。 1あるいは2とMCM1が複合体を形成して、a型および型のいずれの接合型においても細胞特異的な遺伝子の発現を制御している。map1遺伝子産物もまたMCM1と同様に接合型遺伝子産物と相互作用し、P型とM型のそれぞれに特異的な遺伝子の上流配列に結合し、その発現を制御していると申請者は考えている。その際興味深いことに、map1遺伝子はMCM1遺伝子と異なり、細胞分化に特異的に作用し細胞増殖には不要である。またその機能はP型細胞では必須であるが、M型細胞においては絶対に必要なわけでなく、その存在意義は二種の細胞で必ずしも同等ではない。 申請者は出芽酵母MCM1と比較して、map1遺伝子の機能にもう一つの類似性を指摘している。出芽酵母において接合フェロモン受容の情報は、MAPキナーゼ系を経て伝達され最終的にSTE12のリン酸化にいたるが、MCM1はSTE12と複合体を形成することで、これに応答した転写活性化に関与することが示されている。またヒトのSRFは転写因子Elk-1と複合体を形成してc-fosのプロモーターに結合するが、細胞が血清で刺激されるとMAPキナーゼによりElk-1がリン酸化されて転写の活性化がおこる。申請者が明らかにした分裂酵母の細胞内情報伝達系におけるmap1遺伝子の位置は、これらの状況にきわめて類似している。すなわちmap1遺伝子はsxa2遺伝子の転写誘導に関与する因子として単離されており、sxa2の発現はMAPキナーゼ系を介したP-ファクター受容のシグナルにより誘導される。従って、Map1遺伝子も未だ同定されていないMAPキナーゼSpk1の標的タンパク質とともに作用して、接合フェロモン受容系と密接に関与している可能性が高いと申請者は指摘している。 本研究は分裂酵母における細胞型特異的な遺伝子発現の分子基盤を与えている重要な遺伝子を明らかにしたもので、しかも解明された機構は出芽酵母のそれと多くの点で共通点をもっていた。このことはこうした機構が生物種を超えて広く保存されている可能性をも示唆している。今後map1遺伝子産物が制御する、細胞型特異的な遺伝子の上流に共通に存在する配列の同定やMap1と相互作用する因子の同定なども本研究の結果射程距離に入ってきており、これらを通じて分裂酵母の細胞分化の制御機構の全体像が明確なものとなることが期待される。 以上、申請者の研究成果は、分裂酵母の細胞分化の制御機構の理解に大きな進展をもたらしたものであり、申請者は博士(理学)の称号を得るに値するものと委員会は全員一致で判断した。なお、共同研究としてなされた部分について、申請者が最も主要に寄与していることを確認済みである。 |  map1細胞の表現型を詳細に観察した。その結果h-型細胞においてもmap1遺伝子が破壊されると接合能が若干低下することが判明した。次にh-
map1細胞の表現型を詳細に観察した。その結果h-型細胞においてもmap1遺伝子が破壊されると接合能が若干低下することが判明した。次にh-