| 内容要旨 | | 序章 免疫系は自己と非自己を識別し,予測不能な多様性をもつ外界の異物から生体を防御する機構であり,T細胞はその免疫系調節において中心的な役割を担っている。T細胞は抗原レセプター遺伝子の再構成により多様性を獲得するが,T細胞成熟の過程ではその中から自己反応性のものが除去される(ネガティプセレクション)一方,自己のMHCをモチーフにして異物排除に有用なもののみが選別される(ポジティプセレクション)という機構が存在していることが知られている。この2つの選別によるT細胞のレパトワ形成は自己と非自己を識別する上で重要な基礎となっているにもかかわらず,未だ不明な部分が多い。本研究では,まず第1章において,現在のポジティプセレクションのモデルでは説明されないCD8陽性クラスII MHC特異的T細胞の由来について解析し,第2章では,"1抗原レセプター/1リンバ球"の原則を保証していると考えられてきた対立形質排除機構について,特にTCR 鎖に関する解析を行った。第3章では,成熟T細胞の抗原認識におけるco-receptor分子の機能について検討した。 鎖に関する解析を行った。第3章では,成熟T細胞の抗原認識におけるco-receptor分子の機能について検討した。 第1章CD8陽性クラスII MHC特異的T細胞の成熟過程について T細胞は細胞表層上の抗原レセプター(T cell antigen receptor;TCR)により抗原提示細胞上の抗原提示分子(MHC分子)と,それにより提示された抗原ペプチド断片の複合体を認識する。抗原提示分子であるMHC分子は,その構造と機能により2つのクラスに大別され,成熟T細胞はそのTCRがクラスIMHC特異的がクラスII MHC特異的かで2つのサブセットに分けられる。また,多くの成熟T細胞はco-receptor分子と呼ばれる2つの分子,CD4とCD8のうちどちらか片方のみを発現しているため,これによっても2つのサブセットに分類され得る。これら2つの二分律により,成熟T細胞は合計4つのサブセットに分類されるはずであるが,現在一般的に観察されているのはCD8陽性でクラスIMHC特異的な細胞とCD4陽性クラスII MHC特異的な細胞の2つである。このような関係は,CD8がクラスIMHC分子上に,CD4がクラスII MHC分子上にそれぞれ結合部位を持つことがらその合理性が得られており,T細胞がどのMHC分子により提示された抗原を認識するようになるか(MHC拘束性),またどちらのco-receptor分子を発現する細胞に分化するかは,T細胞が成熟する際に受けるポジティプセレクションにより決定されると考えられている。しかしながら,異系リンパ球混合培養により,アロクラスIIMHC特異的な細胞傷害性T細胞(Cytotoxic Tlymphocyte;CTL)を誘導した場合にはCD4陽性細胞のみならず,CD8陽性細胞も同程度かそれ以上の割合で観察される。他方の,アロクラスIMHC特異的なものについてはCD4陽性細胞はほとんど検出されないことから,CD8陽性細胞のアロクラスIIMHCへの反応性は単なる交叉反応ではない可能性が考えられていたが,これらの細胞の成熟機構については未だ説明されていない。そこで本研究では,CD8陽性アロクラスIIMHC(I-Ak)特異的CTLクローン,QM11のTCRトランスジェニックマウスを遺伝学的に解析することにより,QM11TCRを発現した細胞がどのMHC分子によりポジティプセレクションを受けるかの検討を行った。 トランスジェニックマウスは鈴木らにより作製され,1匹のファウンダー(C57BL/6)が得られた。このマウスをQM11が得られたB10.QBRマウス(H-2bq4;Kb,Ib,Dq)と掛け合わせ,脾臓中のT細胞を,QM11TCRのイディオタイプに対する抗体,ID11を用いて解析した。トランスジェニックマウスの脾臓T細胞の中には,CD4陽性細胞,CD8陽性細胞の両方が存在し,また正常マウス中にはほとんど見られないCD4/CD8両陰性の細胞が多数観察された。イディオタイプの発現については,様々な発現量の細胞が観察されたが,CD8陽性でイディオタイプ高陽性の細胞がQM11に相当する細胞群であることが明らかとなった。このCD8陽性,イディオタイプ高陽性の細胞が,どのMHC分子によりポジティブセレクションを受けるかを決定するために,トランスジェニックマウスを様々な系統のマウスと掛け合わせ,その末梢のT細胞の解析を行ったところ,CD8陽性,イディオタイプ高陽性の細胞群はH-2Dq分子を持つマウスにおいてのみ成熟が見られた。すなわち,QM11はクラスIMHC分子,Dqによりポジティプセレクションを受ける細胞であることが明らかとなった。以上の結果より,CD8陽性アロクラスIIMHC特異的T細胞は,自己のクラスIMHC分子によりポジティプセレクションを受けて成熟するという通常のCD8陽性細胞であり,アロクラスIIMHCへの反応性は交叉反応の結果であることが強く示唆された。 第2章T細胞抗原レセプター鎖における対立形質排除について リンパ球上の抗原レセプターは厳密な対立形質排除の機構により1つの細胞上に2種類以上のレセプターが発現することが防がれていると考えられてきたが,QM11TCRトランスジェニックマウスの解析結果はこの原則が必ずしも成立しないことを示していた。ここではTCR 鎖における対立形質排除についてトランスジェニックマウスで得られた知見に基づき正常マウスでの検討も行った。 (1)QM11TCRトランスジェニックマウスにおける解析 QM11TCRトランスジェニックマウスにおいては,末梢の成熟T細胞の約80%が導入遺伝子由来のTCR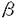 鎖を発現しており,導入遺伝子を発現した細胞内では内在性の鎖遺伝子の再構成,発現は抑制されていた。しかしながらTCR鎖に関しては鎖の場合とは大きく異なり,QM11TCRのイディオタイプ陽性の細胞は10%から30%程度で,しかもトランスジェニックマウス中に多数観察されたイディオタイプ低陽性の細胞は,以下に記す理由により,内在性の鎖を同時に発現したTCR鎖二重発現細胞であると考えられた。 鎖を発現しており,導入遺伝子を発現した細胞内では内在性の鎖遺伝子の再構成,発現は抑制されていた。しかしながらTCR鎖に関しては鎖の場合とは大きく異なり,QM11TCRのイディオタイプ陽性の細胞は10%から30%程度で,しかもトランスジェニックマウス中に多数観察されたイディオタイプ低陽性の細胞は,以下に記す理由により,内在性の鎖を同時に発現したTCR鎖二重発現細胞であると考えられた。 (1)イディオタイプ低陽性細胞は,CD4陽性,CD8陽性どちらの細胞群の中にも存在し,また,どのH-2ハプロタイプ下でも成熟可能であった。このことは,これらイディオタイプ低陽性細胞はQM11TCRとは無関係に成熟していることを示唆していると思われる。(2)これらの細胞は,イディオタイプの発現は低いが,TCR 鎖の発現量は正常である。(3)QM11TCRの鎖にはVA10が用いられているが,トランスジェニックマウスの脾臓T細胞を抗イディオタイプ抗体とV3.2,V8,V11に対する抗体のmixtureとで二重染色を行ったところ,イディオタイプ低陽性の細胞群の中にはV(3.2+8+11)を発現した細胞が含まれていた。(4)トランスジェニックマウスを,遺伝子再構成能に欠陥を持つSCIDマウスと掛け合わせることにより,内在性のTCR遺伝子の発現を抑制した場合にはイディオタイプ低陽性細胞の成熟は見られなくなった。 実際,トランスジェニックマウスの脾臓T細胞を抗イディオタイプ抗体と抗V3.2抗体とで交互に刺激することによって,イディオタイプとV3.2の両方を同時に発現した細胞株を樹立することができた。 (2)正常マウスからのTCR鎖二重発現細胞株の樹立 QM11TCRトランスジェニックマウスで多数観察されたTCR鎖二重発現細胞が,真に正常マウスにも見られる事象を反映しているのかを検討するために,つぎに正常マウスにおける解析を行った。 Vはその数も多く(〜100個程度),正常マウスでは特定の組み合わせのV二重発現細胞はもし存在していても頻度が低いことによりフローサイトメトリーでは検出できないことが考えられたため,TCR鎖二重発現細胞の株化を試みた。正常C57BL/6マウスの脾臓T細胞をまずV3.2に対する抗体で刺激し,すべての細胞がV3.2陽性であるような細胞株を得た。その後,その一部については抗V8抗体で2週間おきに3回刺激を繰り返したところ,V3.2,V8どちらについても陽性であるような細胞株が得られ,正常マウス中にもTCR 鎖二重発現細胞が存在することが明らかとなった。この細胞株の中にはV3.2/V8の発現量比の異なるいくつかの細胞群が含まれていたが,この量比は1年以上in vitroで継代した後でも変化せず,非常に安定であった。また,TCR二重発現細胞上の2つのTCRは,どちらから刺激を加えてもT細胞に細胞傷害,顆粒放出,増殖などの反応を誘起することができ,どちらも機能的であることが示された。さらにTCR 鎖二重発現細胞はこの組み合わせ以外にV2/V3.2,V2/V8の組み合わせのものも,別の系統のマウスから樹立することができた。これらの結果は,TCR 鎖の対立形質排除は成立していないことを示唆していると考えられる。 第3章成熟T細胞の抗原認識,活性化における co-receptor の機能の解析 co-receptor分子はシグナル伝達分子としての機能を持つと考えられているものの,その具体的な機能としてはSrc系のプロテインチロシンキナーゼp56lckを介したもの以外には知られていない。成熟T細胞は抗原を認識し,活性化するとさまざまな反応を示すが,本研究ではco-receptorを介したシグナルがそのいずれかの反応に関与しているか否かを検討する目的で以下の実験を行った。 (1)co-receptor 欠損変異株の取得 アロクラスII MHC(I-Ak)特異的なQM11の細胞傷害活性は抗CD8抗体での阻害を受けず,QM11の抗原認識はCD8分子の細胞外部分の機能(MHC結合補佐機能)は必要なく,TCR/MHCの相互作用のみで成立すると考えられる。このことはQM11TCRトランスジェニックマウスから樹立されたCD4陽性でQM11TCR陽性のT細胞クローン,4Q11の抗原認識も抗CD4抗体で阻害を受けないことからも示唆された。QM11TCR陽性のT細胞クローン,4Q11の抗原認識も抗CD4抗体で阻害を受けないことからも示唆された。そこで,QM11や4Q11からそれぞれCD8,CD4の欠損株8N11,4N11を樹立し,その抗原認識後の反応性について親株と比較することにより,T細胞活性化におけるco-receptor分子のシグナル伝達機能の解析を行った。 (2)co-receptor 欠損変異株の反応性についての検討 T細胞活性化後の反応におけるco-receptor分子の機能について解析するために,QM11と8N11については細胞傷害,顆粒放出, -IFNの産生,および増殖の4つの反応に関して,4Q11と4N11については細胞傷害,T細胞増殖因子の産生,増殖の3つの反応に関してそれぞれ比較,検討した。その結果,CD8,CD4どちらの欠損株についても,すべての反応において抗原特異的な,親株と同程度の活性が見られ,co-receptor分子,およびco-receptorを介したシグナルはT細胞の活性化には不可欠ではないことが強く示唆された。 -IFNの産生,および増殖の4つの反応に関して,4Q11と4N11については細胞傷害,T細胞増殖因子の産生,増殖の3つの反応に関してそれぞれ比較,検討した。その結果,CD8,CD4どちらの欠損株についても,すべての反応において抗原特異的な,親株と同程度の活性が見られ,co-receptor分子,およびco-receptorを介したシグナルはT細胞の活性化には不可欠ではないことが強く示唆された。 (3)co-receptor 欠損株の活性化におけるp56lckの関与について T細胞の活性化は,co-receptor分子がTCRと同じMHC分子と結合することによりco-receptorの細胞質内部分に会合しているp56lckがTCR複合体中の 鎖に接近し,そのチロシン残基をリン酸化することによって開始されると考えられている。そこでここではこのリン酸化がco-receptorがない場合にも起こり得るかの検討を行った。その結果,co-receptor欠損株においても抗原特異的に鎖のリン酸化が観察され,p56lckはco-receptor分子に会合していなくてもT細胞活性化に関与しうることが示唆された。 鎖に接近し,そのチロシン残基をリン酸化することによって開始されると考えられている。そこでここではこのリン酸化がco-receptorがない場合にも起こり得るかの検討を行った。その結果,co-receptor欠損株においても抗原特異的に鎖のリン酸化が観察され,p56lckはco-receptor分子に会合していなくてもT細胞活性化に関与しうることが示唆された。 (4)抗原提示細胞上の抗原量とco-receptor分子への依存性について QM11や4Q11の抗原認識において,抗原提示細胞上の抗原の量が多い場合にはco-receptor分子の寄与は見られないが,抗原の量が減ってTCR/MHCの相互作用のみでは十分な親和活性が得られなくなった状況下ではco-receptor分子が大きく関与することが考えられた。実際,抗I-Ak抗体を適量用いることにより利用可能なI-Ak分子の量を変えてQM11と4Q11の細胞傷害活性を測定すると,4Q11については高濃度の抗I-Ak抗体存在下(利用可能なI-Ak量が相対的に少ない場合)でも抗原提示細胞を認識し,作用を及ぼすことが示された。ただしその場合,抗CD4抗体による阻害が見られ,CD4依存的に抗原認識を行っていると考えられる。CD4に対する依存性は抗原量が少なくなる程増していった。一方,co-receptor分子がTCRと同一のMHCと結合することのできないQM11の場合には,抗I-Ak抗体による阻害を受けやすく,また抗原量が減少していっても,CD8分子による寄与はほとんど観察されなかった。以上の結果より,co-receptor分子は抗原量が少ない場合においてもT細胞の抗原認識を成立させるという機能を果たしており,そのような効果はco-receptorとTCRが同じMHC分子と結合できる場合にのみ顕著に発揮されることが示唆された。生理的条件下では,1種類の抗原ペプチドがMHC分子によって提示される量は限られていると考えられ,このような場面においてはco-receptor分子はT細胞の抗原認識に不可欠であると考えられる。 |
 は数が多く、単に末梢のT細胞を特定の2つの抗
は数が多く、単に末梢のT細胞を特定の2つの抗