高等植物は香味料や薬品に使用されるそれぞれの種に特異的な有機化合物を多く含んでいる。このような有機化合物は植物生理的には二次代謝産物として知られている。しかし、未だそれらの物質の多様性や蓄積条件などを完全に解明するには至ってはいない。 フェニルプロパノイド化合物代謝経路は、フェニルアラニンを出発物質とする二次代謝経路で、クマリン、フラボノイド、リグニンなどを代謝産物としている。この代謝系に関与する酵素群がどのように誘導されるかは明らかになりつつあるが、この経路全体の制御関係は未だに解明されていない。その解明手段の一つとして、フェニルプロパノイド経路上流の鍵酵素フェニルアラニンアンモニアリアーゼ(PAL)遺伝子を導入した植物を作製し、PAL活性の上昇によるフェニルプロパノイド経路への影響の解析が挙げられる。現在まで、代謝経路上流に位置する酵素の遺伝子を外部より追加したトランスジェニック植物の代謝産物変化についてはほとんど報告されていない。さらに、その代謝経路全体への影響はほとんど知られておらず、遺伝子導入による植物の二次代謝や生理の変化についての知見はこれからの課題となっている。 本研究では、辛味成分として知られ、フェニルプロパノイド経路の行き止まり化合物であるカプサイシンを生産するトウガラシ(Capsicum trutescens)を材料として用いた。筆者らは、まず安定な培養細胞系を確立することを試みた。次いで、それにPAL遺伝子を導入し、代謝の流れの一部を太くした場合にそれが二次代謝の流れにどう影響を及ぼすのかを検討した。さらに、同じ系を用い、物質的な刺激がどのような影響を及ぼすかも検討した。 1.トウガラシ毛状根培養系の作製 植物細胞の遺伝的な操作による代謝変更と刺激に対する二次代謝応答の影響を比較して検討するためには、できるだけ均一な条件の細胞が必要とされる。この条件に当てはまる培養細胞系として、毛状根培養を選択して研究を進行した。 毛状根は、Capsicum frutescens胚軸にAgrobacterium rhizogenesを接種することにより得た。3種のA.rhizogenes A5,A13,1610株を用いたが、それらの菌株によって毛状根の発生率は異なっていた。またそれぞれによって誘導された毛状根には、形態及び成長速度に違いがあった。1610株の接種からは、安定に増殖するtumorも得られた。対照としての通常植物体由来の根の培養も試みたが、継続的な成長は得られなかった。 A13株接種から得られた毛状根における、Riプラスミド上のT-DNAの感染を検証するために、PCRによるT-DNA内のrol遺伝子の検出を行った。A.rhizogenes A13,1610株誘導の安定して成長する毛状根株から、目的遺伝子に相当するPCR産物を検出した。この産物はサザン・プロットによりrol遺伝子プローブとハイブリダイズした。 トウガラシ実生の根、茎、葉は、カプサイシンのフェニルプロパノイド前駆体及びカプサイシンを蓄積せず、A13株由来毛状根もカプサイシンを生産しなかった。 2.PAL遺伝子形質転換体の作製 PAL遺伝子形質転換体作製のための遺伝子導入は、バイナリーベクターを持つA.rhizogenesを用い、毛状根の発生時の形質転換として行った。バイナリーベクターには、リポーター遺伝子として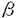 -グルクロニダーゼ(GUS)遺伝子を持ち、プロモーターとしてCaMV35Sを持つpBI121と、そのGUS遺伝子をパセリ由来のPAL cDNAに置換したpBI12PPALを用いた。コントロールとして、GUS遺伝子の形質転換体も同時に作製した。使用したバイナリーベクターは、形質転換選択マーカー遺伝子としてカナマイシン代謝酵素NPTIIを含んでいるが、寒天培地上での毛状根の生育はきわめて不安定であったため、液体培地に移行させることができたクローンしかカナマイシンによる選択が行えなかった。 -グルクロニダーゼ(GUS)遺伝子を持ち、プロモーターとしてCaMV35Sを持つpBI121と、そのGUS遺伝子をパセリ由来のPAL cDNAに置換したpBI12PPALを用いた。コントロールとして、GUS遺伝子の形質転換体も同時に作製した。使用したバイナリーベクターは、形質転換選択マーカー遺伝子としてカナマイシン代謝酵素NPTIIを含んでいるが、寒天培地上での毛状根の生育はきわめて不安定であったため、液体培地に移行させることができたクローンしかカナマイシンによる選択が行えなかった。 バイナリーベクターによる遺伝子導入及びその発現は、GUSのヒストケミカル・アッセイによって確認した。GUS遺伝子をターゲットとしたPCRによる検出も行い、GUS活性を示したクローンは、全てPCRによるバンドの増幅を示した。 次に、カナマイシンによって選択された、pBI12PPAL保持A.rhizogenes接種より得られた毛状根へのNPT II領域のPCRを行ったところ、全てのクローンで、相当するバンドの増幅を検出した。さらに、PAL遺伝子領域をプローブとしたサザン・ブロットによってpBI12PPAL形質転換体の確認を行った。この遺伝子の転写は、ノーザン・ブロットによって確認した。 これらのpBI12PPAL形質転換体は、いずれも褐色化し、根の直径が通常の3倍程度に増大するという形態的特長を示した。また、コントロール毛状根、pBI121形質転換体と比較して、極端に成長が遅く、成長に対応したPAL活性も全く異なるパターンをとっていた。これに対して、pBI121形質転換体の形態は、コントロールとなる毛状根と全く変化がなかった。 3.PAL遺伝子形質転換体における生理的変異とフェニルプロパノイド化合物代謝の解析 前述の3種の遺伝子型(コントロール毛状根、pBI121及びpBI12PPAL形質転換体)の解析を行った。 湿重量あたりの可溶性タンパク量及び粗脂肪量は、3種の遺伝子型でいずれも同様の値を示した。pBI12PPAL形質転換体は遊離アミノ酸量について、ほとんどのアミノ酸をコントロール毛状根より多く含んでいた。pBI12PPAL形質転換体の湿重量あたりの乾燥重量は、コントロール毛状根、pBI121形質転換体の約2倍であった。2種類の粗繊維の含量の測定は、pBI12PPAL形質転換体においてリグニン関連物質が多く蓄積していることを示し、これは乾燥重量の約60%を占めていた。また、クロロホルム可溶物質と酸処理によってクロロホルム可溶となる物質の中で、pBI12PPAL形質転換体に特異的に蓄積する物質が存在していた。 フェニルプロパノイド代謝産物については、フェルラ酸と、リグニン様物質がpBI12PPAL形質転換体においてより多く蓄積をされていた。しかしながら、フェルラ酸を除いたカプサイシン前駆体及びカプサイシンの大きな蓄積は検出されなかった。 PAL遺伝子の導入がフェニルプロパノイド化合物代謝に及ぼす影響を、さらに解析するために、トウガラシ毛状根へ物質的な刺激を与え、その影響を検討した。酵母抽出物、グルタチオン、硫酸銅、キトサン、過酸化水素、サリチル酸、フェニルアラニン、8-methylnonanoic acidの各物質を、それぞれ異なる刺激を細胞に与えるために培地に添加した。これらの刺激物質の処理からフェニルプロパノイド化合物代謝については、以下の4点が明らかになった。1.フェニルアラニンと8-methylnonanoic acid以外の処理は、毛状根組織へのフェニルプロパノイド化合物蓄積にほとんど影響がない。2.全ての処理が、培地中へのフェニルプロパノイド化合物の漏出を誘導する。3.培地へのフェニルプロパノイド化合物漏出のパターンは、処理によって異なっている。4.毛状根が褐色化した処理では、リグニン様物質含量が増加する。 リグニン化を最も激しく引き起こしたサリチル酸処理とフェニルプロパノイド化合物代謝全体の基質となるフェニルアラニン処理の毛状根のフェニルプロパノイド代謝応答の際の、鍵酵素PALとカプサイシン合成酵素について活性を測定した。その結果、それぞれの酵素は量的、時間的に異なる誘導を受けていることが分かった。また、リグニン様物質含量の増加は、PAL誘導から遅れて生じているため、桂皮酸からリグニンへの代謝が逐次的ではないことが示された。 pBI12PPAL形質転換体への刺激物質サリチル酸とフェニルアラニンの処理は、コントロール毛状根やGUS遺伝子導入形質転換体と同様に、フェニルアラニンによるわずかな成長減少及びサリチル酸による生育の停止を引き起こした。クロロホルム可溶物質および酸処理後のクロロホルム可溶物質については、全ての遺伝子型についてサリチル酸処理による物質の減少が観察されたが、pBI12PPAL形質転換体では刺激物質によってリグニン様物質量は増加しなかった。 以上、トウガラシ毛状根細胞において、PALの遺伝子導入と刺激物質とがフェニルプロパノイド代謝にもたらす影響を検討した。その結果、以下のことが明らかになった。1.PAL遺伝子の導入によって、フェニルプロパノイド化合物代謝は、特にカプサイシンの生合成へは向かわず、行き止まり化合物のリグニン様物質の生合成が生育限界まで進行する。2.カプサイシン前駆体としては、フェルラ酸、バニリンが組織中に蓄積し、桂皮酸、クマル酸、カフェー酸は蓄積しない。3.培地へのカプサイシン前駆体の漏出は、物質添加による刺激によって生ずるが、pBI12PPAL形質転換体では引き起こされなかったため、代謝の亢進が必ずしも組織外への漏出とつながらない。 これより、カプサイシン生合成の誘導は、フェニルプロパノイド化合物代謝上流遺伝子の形質転換によっては行えず、今後、カプサイシン合成酵素及びその遺伝子を取得することにより、形質転換系を用いたカプサイシン生合成の操作が期待される。 |