5-Azacytidine(5Az)はcytidineのアナログで、抗癌剤として開発されたが、副作用も多く、とくに神経系、心臓、骨格筋に対する毒性が知られている。また、催奇形性も有しており、妊娠マウスに投与すると胎仔の神経系に異常を引き起こす。近年、5Azが10T1/2や3T3などマウスの培養間葉細胞を分化させることが明らかになった。この作用は、5AzがDNAのメチル化を抑制し、非活性状態にあった各種遺伝子の発現を誘導するためと考えられている。 一方、アポトーシスは近年提唱された新しい細胞死の概念で、予め遺伝子にプログラムされた死への機序が生理的または病理的に活性化されたときに生じる。この型の細胞死は核DNAの断片化が細胞質の変化に先行し、細胞核の凝縮、断片化を形態的特徴とする。最近では様々な化学物質によって誘発されたアポトーシスの報告が増加している。 我々は5Azの飲水投与を続けているマウスの家系に無または小眼症の仔が生じることを見出した。そして、この変化が発生期における神経系の細胞死亢進によるもので、さらにこの細胞死は5AzによるDNA脱メチル化のために何らかのアポトーシス関連遺伝子が活性化したために生じたと推定した。 この仮説を証明するためには5Az誘発による神経系細胞アポトーシスの実験系が是非とも必要と考えた。そこで、この実験系の確立を目的として以下のような研究を行った。 最初に、神経系に対する5Azの催奇形性を明らかにするため、妊娠マウスに5Azを単回腹腔内投与し、胎仔の生死および誘発病変の性質とその推移をしらべた。ICR、C57BL/6、BALB/cの妊娠マウスを用い、妊娠11日目に5Az(0.2〜0.4mg)を腹腔内投与し、胎仔の生死を観察した。また妊娠11日のICRマウスに5Azを投与後、経日的に胎仔を採取、光顕および電顕的に観察した。 5Az投与後7日(妊娠18日)に胎仔を観察したところ、死亡して吸収胚となっているものと正常に発育しているものとがあった。吸収胚の割合は、ICRでは5Az0.2mg/head投与で30.4%、0.4mgで71.2%、C57BL/6では0.2mgで67.2%、BALB/cでは0.2mgで53.2%であった。ICRマウスを用いた病理組織学的検索では、投与1日後に胎仔の脳と脊髄に広範な細胞死領域が認められた。病変部の細胞は核が濃縮し、粉砕されているものが多かった。破壊された核は近接する細胞やマクロファージに貪食されていた。この組織を切片上でアポトーシスを検出するTUNEL法で染色したところ、上述の濃縮・粉砕核が陽性を示した。電顕観察ではこれらの細胞に核クロマチンの濃縮と核辺縁への凝集、核の断片化を認め、細胞質では空胞の形成、濃縮、縮小等がみられた。断片化した核は周辺の細胞およびマクロファージに貪食されていた。このような神経組織の細胞死病変は5Az投与後2日で軽減し、3日後にはほとんど正常の組織に戻った。 以上のことから、5Azは妊娠中期胎仔の主として中枢神経組織に細胞死を誘発すること、この細胞死は形態学的にアポトーシスであることが示された。病変の軽い胎仔では、神経組織は残存する未分化細胞の分裂によって速やかに補填され、胚の正常発育が続くと考えられた。 次に、マウス胎仔の神経系初代培養細胞における5Azの作用を調べた。妊娠11日目のICRマウス胎仔の全頭部を細片にし、トリプシンとコラゲナーゼで処理し、細胞浮遊液を作った。これを1週間培養したところ、間葉細胞がシート状に広がり、その上に長い突起を有する三角形または紡錘形の神経様細胞が散在する混合細胞系が得られた。これに5Az(0,5,50,500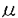 g/ml)を加え、翌日に細胞を固定、光顕、走査電顕および透過電顕で観察した。アポトーシスの確認はTUNEL法で行った。 g/ml)を加え、翌日に細胞を固定、光顕、走査電顕および透過電顕で観察した。アポトーシスの確認はTUNEL法で行った。 5Az非投与群(0g/ml)では間葉細胞がシート状に広がり、この上にBodian法で染色される長い突起を有する神経様細胞が散見された。走査電顕でも間葉細胞のシート状増殖と突起を有する神経様細胞が確認された。5g/ml投与群では間葉細胞は変化を示さなかったが、神経様細胞の細胞体は濃縮し、突起は消失していた。走査および透過電顕観察では、核クロマチンの濃縮、凝集が、一部では小型円形の核断片(アポトーシス小体)の集合が認められた。TUNEL法でアポトーシスの確認を行ったところ、濃縮核または崩壊した核質の凝集体に一致して陽性像が観察された。50および500g/ml投与群では神経細胞は全く認められず、間葉細胞も広範な細胞崩壊を示したが、これは細胞質の崩壊を主とする壊死(Necrosis)であると考えられた。 今回の実験から、5Azでアポトーシスが誘導される細胞(神経細胞)と誘導されない細胞(間葉細胞)とがあり、前者では低量でアポトーシスが誘導され、後者では多量投与によって壊死が誘導されることがわかった。この結果、発生期の神経細胞が5Azによるアポトーシス誘導の標的細胞であることが確認された。 株化神経細胞での5Azの作用を調べるため、神経系細胞のin vitroモデルとして多用されているPC12細胞を用いた。神経成長因子(NGF)未添加の未分化PC12およびNGFを添加して神経様細胞に分化したPC12に5Az(0,5,50,100,500g/ml)を加え、30時間後の形態変化を光顕、走査電顕および透過電顕で観察した。アポトーシスの確認はTUNEL法で行った。 この結果、5Azは未分化PC12、NGF処置分化PC12のいずれにもアポトーシスを誘導した。アポトーシスを起こしたPC12細胞は形態的には前述の初代培養神経細胞と同様の変化を示した。すなわち、細胞核の濃縮、断片化およびアポトーシス小体の形成などである。これらの細胞核はTUNEL法陽性であった。このアポトーシスは5Az用量依存性であったが、未分化細胞が50g/mlでアポトーシスを起こしたのに対し、分化細胞では100g/mlでようやく明瞭なアポトーシスが認められ、未分化細胞の方が感受性が高かった。すなわち、5Azは株化神経様細胞にもアポトーシスを誘導すること、および細胞分化の程度によって5Azに対する感受性が異なることが明らかになった。 次に、上述した5Az誘発PC12細胞アポトーシス実験系の生化学的性質を調べた。まず、DNAの断片化を確認するために、アポトーシス細胞のDNAを電気泳動した。次にこのアポトーシスの発生機序を明らかにするため、分化PC12細胞を蛋白合成阻害剤cycloheximideまたは神経系細胞のアポトーシスを抑制すると報告されている各種化学物質(cAMP,KCl,deoxycytidine)で処理し、5Az誘発アポトーシスの抑制の有無を調べた。細胞死の程度は培地へのLDH放出量でモニターした。LDH放出量は細胞質崩壊の指標で、形態的なアポトーシスの出現から24ないし48時間遅れて現れた。 DNA電気泳動の結果、5Az処理した未分化および分化PC12細胞に生化学的特徴であるDNAの断片化を示すラダー形成が認められた。また、分化PC12細胞の5Az誘発アポトーシスはcycloheximideとcAMP処置によって対照レベル近くまで抑制されたが、KClまたはdeoxycytidine処理では抑制されなかった。 以上のことから、5AzによるPC12細胞アポトーシスの誘導には新たな蛋白質の合成が必要であり、さらにこの蛋白合成系にはcAMPを介するシグナル伝達機構が存在することが示唆された。 本研究の結果から、5Azは妊娠中期の胎仔神経系細胞、株化神経様細胞など発生増殖期の神経系細胞にアポトーシスを誘発することが明らかになった。さらに、この5Az誘発アポトーシスには新たに合成された未知のアポトーシス関連蛋白質の関与が推定された。この蛋白質が5Azの脱メチル化作用によって発現したものかどうかは今後解明すべき課題と思われる。本研究で確立された実験系はこの課題の解明に不可欠であり、神経系におけるアポトーシス研究一般においても有用となろう。 |