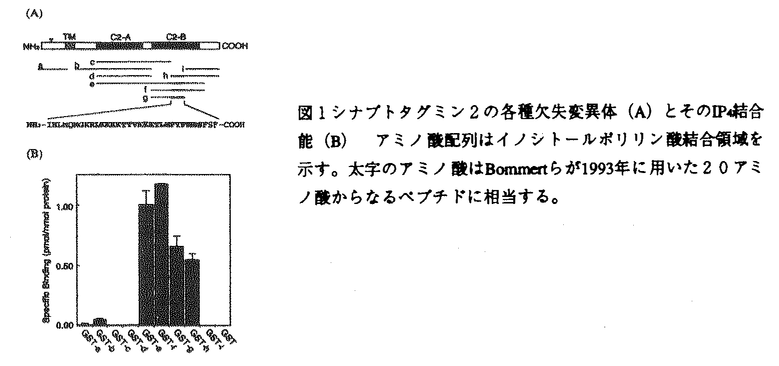
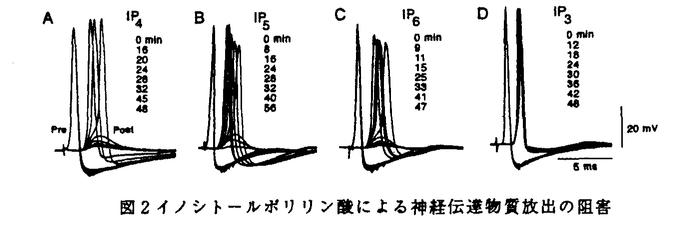
本研究は細胞内情報伝達に重要な役割を演じていると考えられているイノシトールポリリン酸(イノシトール1,3,4,5-四リン酸(IP4)、イノシトール1,3,4,5,6-五リン酸(IP5)、イノシトール1,2,3,4,5,6-六リン酸(IP6))の生理的機能を明らかにするため、マウス小脳よりこれらの結合蛋白質の分離・精製試みたものであり、下記の結果を得ている。 1.可溶化したマウス小脳膜画分から[3H]IP4の結合活性を指標として一連のカラムクロマトグラフィーによりイノシトールポリリン酸結合蛋白質を分離・精製した。部分アミノ酸配列をもとにマウス小脳cDNAライブラリーを検索した結果、この蛋白質はシナプス小胞に多量に存在し、神経伝達物質の放出に重要とされているシナプトタグミン2と同定された。 2.シナプトタグミンはN末端側に膜貫通領域を一カ所持ち、C末端側の細胞質内領域にはプロテインキナーゼのC2調節領域とホモロジーを有する配列が二回繰り返しておりそれぞれC2A,C2Bドメインと呼ばれている。これらのドメイン構造をもとに様々なデリーションミュータントをグルタチオンS-トランスフェラーゼとの融合タンパク質として発現させイノシトールポリリン酸結合部位についで検討したところ、イノシトールポリリン酸はC2Bドメインの中央約30アミノ酸にのみ結合し、C2Aドメインには全く結合しなかった。このイノシトールポリリン酸結合部位は神経伝達物質の放出に重要とされている領域とほぼ一致することからイノシトールポリリン酸の神経伝達物質放出の調節因子としての機能が示唆された。 3.IP4(IP5,IP6)をイカの巨大軸索のプレシナプスへマイクロインジェクションしたところ、IP4(IP5IP6)はプレシナプスの活動電位及びCa2+電流には何ら影響を与えずに、神経伝達物質の放出を抑制することが明らかとなった。また比較としてIP3をインジェクションしても、このような抑制効果は全く認められなかった。 4.C2Aドメインに対する抗体をイカの巨大軸索のプレシナプスに導入したところ、イノシトールポリリン酸の場合と同様な神経伝達物質放出の阻害が観測された。電子顕微鏡による観察からC2A抗体をインジェクションしたプレシナプスではシナプス小胞の数がコントロールに比べ30%増大しており、シナプス小胞の融合の過程が阻害されているものと考えられた。 5.C2Bドメインに対する抗体をイカの巨大軸索のプレシナプスに導入したところ、プレ及びポストの活動電位に何ら変化は認められなかったが、IP4と同時にインジェクションすることによりIP4の神経伝達物質放出の阻害効果を完全に回復することが示された。またC2B抗体を導入したプレシナプスに高頻度刺激を加えその電子顕微鏡像を観察したところ、シナプス小胞の数がコントロールに比べ90%近く減少しており、シナプス小胞のリサイクルがシナプトタグミン分子のC2Bドメインを介して行われていることが示唆された。 以上、本論文はシナプトタグミン分子のイノシトールポリリン酸結合領域の生化学的解析、及びイカ巨大軸索のプレシナプスへのマイクロインジェクションを用いた生理学的解析から、イノシトールポリリン酸が神経伝達物質放出の負の制御因子である可能性を明らかにした。本研究はこれまで未知に等しかった、イノシトールポリリン酸の生理学的機能の解明に重要な貢献をなすと考えられ、学位の授与に値するものと考えられる。 |