I.ヒトGM-CSF受容体遺伝子の構造と発現の解析 1.血液系のうち主に骨髄系の細胞の増殖・分化を制御するGM-CSFの受容体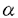 鎖(hGMR)遺伝子の全長をクローニングし、そのエクソン・イントロン構造および転写の開始部位を明らかにした。hGMR遺伝子の全長は約44kbで13のエクソンからなっていた。また、プライマー伸長法と5’RACE(Rapid amplification of cDNA end)法を行ったところ、どちらの方法においても見いだされる主要な転写の開始部位を一つ同定した。
鎖(hGMR)遺伝子の全長をクローニングし、そのエクソン・イントロン構造および転写の開始部位を明らかにした。hGMR遺伝子の全長は約44kbで13のエクソンからなっていた。また、プライマー伸長法と5’RACE(Rapid amplification of cDNA end)法を行ったところ、どちらの方法においても見いだされる主要な転写の開始部位を一つ同定した。
2.hGMR遺伝子のエクソン・イントロン構造はhGMR蛋白質の構造によく対応していた。すなわちエクソン・イントロンの境界部が細胞外領域に3つ存在するファイブロネクチンタイプIII様ドメイン(Fn3ドメイン)の境界部によく一致し、各ドメインがそれぞれ2つあるいは3つのエクソンによりコードされていた。さらに、エクソン・イントロン境界部のコドンにおけるイントロンの挿入部位を詳細に検討し、他の既知のサイトカイン受容体遺伝子のそれと比較した結果、単一のFn3ドメインをコードする遺伝子断片が重複、挿入を繰り返すことによってすべてのサイトカイン受容体に共通の祖先遺伝子を作り出したという進化上のシナリオが遺伝子構造のなかに明瞭に現れていることを明らかにした。さらに、異なる受容体遺伝子の構造を比較することにより、受容体ファミリーの各メンバーの進化上の関係について一つのモデルを提唱した。
3.転写開始部位の上流約400bpの塩基配列を決定したところ、hGMR遺伝子のプロモーター領域にはTATA boxが存在せず、その代わりにGGGAGGGというプリンに富んだ配列が見いだされた。プロモーター領域内のエレメントとして、転写開始点より45bp上流に、骨髄系細胞、B細胞に特異的な転写因子PU.1の認識配列であるGAGGAA配列が認められた。さらに、Northern解析により成人の各臓器におけるhGMR遺伝子の発現を検討した結果、心臓、脳、胎盤、肺、肝臓、骨格筋、腎臓、膵臓といった多くの非造血組織がこのこの遺伝子を発現していることを明らかにした。
II.中枢神経系幹細胞株を用いた、神経上皮における領域特異的遺伝子の発現制御の解析 1.ラット胎児の前脳、中脳の神経上皮細胞から樹立された中枢神経系幹細胞株(MNS細胞株)を用いて、胎児の神経上皮の特定の領域のみに限局する種々の転写因子の発現がどのように制御されているかを検討した。試験管内でニューロン及びグリアに分化しうる5種類の異なるMNS細胞株について、未分化状態における領域特異的遺伝子の発現を定量RT-PCR法を用いて解析した結果、異なる細胞株が、前脳において領域特異的に発現しているFax-3,Pax-5,Pax-6,Dlx-1,Dlx-2,Otx1,Emx2,Dbxなどの遺伝子をそれぞれ異なった組み合わせで発現していることが明らかとなった。これに対して、後脳よりも後方、あるいは中脳より後方のみに発現している遺伝子であるHox-B1,Hox-B2およびEn-1は、いずれの細胞株においても発現していなかった。この結果は、用いた細胞株が前脳と中脳の神経上皮に由来していることとよく一致していた。
2.これらの細胞株をニューロンおよびグリアへと分化誘導しても、上記の領域特異的遺伝子の選択的発現は基本的に変化しなかった。また、単一細胞レベルでの発現を検討したPax-6の場合は、未分化状態でPax-6を高いレベルで発現している細胞株においては、分化誘導によって発生したMAP2陽性のニューロンにFax-6の発現が受け継がれていることが明らかとなった。このことから、未分化な神経上皮細胞における領域特異的遺伝子の発現(の少なくとも一部)は、その子孫であるニューロンにおける領域特異性を規定していると考えられた。同様の結果は、初代培養した神経上皮細胞でも観察された。上記の実験結果は、神経上皮細胞において自身の領域特異的な性質を維持する何らかの細胞内在的な機構が存在しており、これが領域特異的な遺伝子発現の維持、さらには分化したニューロンやグリアのサブタイプを規定していることを示唆していると考えられる。
3.次に、腹背軸を決定する細胞外シグナルであるSonic hedgehog(SHH)をMNS細胞株に加え、遺伝子発現の変化を解析した。用いた各細胞株は内在的にはSHHを発現していないが、外部からSHHを作用させることにより、胎児脳の腹側の神経上皮あるいはニューロンに発現しているIsl-1,Nkx-2.1,Nkx-2.2の3遺伝子の誘導能の点において、それぞれ異なる応答性を示した。このことから、各細胞株に内在する領域特異性の違いがSHHに対する異なった応答性を生じていることが示唆された。
以上、本論文の前半は、造血の重要な制御因子であるGM-CSFの受容体の発現制御に関する知見を与え、また、造血系が進化の過程でどのようにして、数多くのサイトカイン受容体遺伝子を生じるに至ったかについての新たな考察を行った。後半では、未分化神経上皮における領域特異的遺伝子の発現制御機構を解析する有用な実験系を確立することにより、中枢神経系を構成する多様な細胞がどのような分子機構で生じるかという、重要な生物学上の問題の解明に道を開いた。以上の点より、本論文は学位の授与に価するものと考えられる。
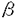 鎖の2つのサブユニットから構成される。これらおよび他のサイトカイン受容体を構成する蛋白質は、細胞外領域にいくつかの共通した構造した構造、1)WSXWSモチーフ、2)通常4つの、ほぼ等間隔で並んだCystein残基、3)通常2つの、約100アミノ酸からなり特徴的な2次構造を持つドメイン(フィブロネクチンのタイプIIIドメインに類似しているため、Fn3ドメインと呼ぶ)がある。申請者は、ヒトGM-CSF受容体
鎖の2つのサブユニットから構成される。これらおよび他のサイトカイン受容体を構成する蛋白質は、細胞外領域にいくつかの共通した構造した構造、1)WSXWSモチーフ、2)通常4つの、ほぼ等間隔で並んだCystein残基、3)通常2つの、約100アミノ酸からなり特徴的な2次構造を持つドメイン(フィブロネクチンのタイプIIIドメインに類似しているため、Fn3ドメインと呼ぶ)がある。申請者は、ヒトGM-CSF受容体