| 背景 神経系を構成する多様な神経細胞が発生の過程でどのように生まれるかという問題は、神経系を理解するうえで重要な問題である。 神経発生研究のモデル生物として、キイロショウジョウバエは以下の点で大変魅力的である。 1.遺伝学的手法により神経発生に関わる未知の遺伝子を同定しそれらの相互作用を解析できる。 2.キイロショウジョウバエで明かにされた形態形成の仕組は、脊椎動物でも機能することが分かっている。神経発生においても普遍的な機構を明かにすることができるものと期待される。 3.腹部中枢神経系の構造が単純なので解析がしやすい。 以上の理由から、私は実験材料としてキイロショウジョウバエを選び、神経発生に必須な遺伝子、prospero(pros)を研究の対象とした。 序論 pros遺伝子の機能喪失変異の胚では腹部中枢神経系に大きな形態異常が生じ、致死となる。遺伝子の解析から、pros遺伝子はPEST配列、グルタミンストレッチ、核移行シグナル、そしてカルボキシル末端(C末)に近いところにホメオドメインを持つ1403アミノ酸からなる転写因子をコードすることが明かになった。 ショウジョウバエでは幹細胞の神経芽細胞が非対称分裂により二次前駆細胞である神経母細胞を次々に生み、更にその神経母細胞がもう一度分裂して、神経細胞が生まれてくる。pros遺伝子は神経芽細胞と神経母細胞の両方の細胞で転写される。ところがProsタンパク質は神経芽細胞の核には認められず神経母細胞の核にのみ局在する。このことから、Prosタンパク質は神経母細胞の段階で一過性に遺伝子発現の調節を行なうことにより、神経細胞の分化を制御するものと考えられ、神経母細胞で様々な遺伝子発現に異常が生じるpros欠失変異の表現型と一致する。 Prosタンパク質は神経芽細胞でも翻訳されるが核には移行せず、細胞周期の進行に伴い神経母細胞に輸送されるという非常にダイナミックな振舞をする。即ち、Prosタンパク質は静止期には細胞質に分布しているが、分裂期に入ると、神経母細胞が発芽する位置の細胞膜の直下に三日月状に集積してくる。その後、anaphase、telophaseを経る過程で集積したProsタンパク質は、神経母細胞の発芽とともに細胞膜直下に分布したまま神経母細胞へ輸送され、細胞質分裂の完了後、神経母細胞の核に移行する。 非対称分配の分子メカニズムの解析 Prosタンパク質が神経芽細胞では核移行を行わず細胞質にとどまり、非対称分裂に伴い神経母細胞に分配されることは、pros遺伝子の機能の発現に重要な意味を持つものと考えられる。この分子機構の解明を目指して解析を行ない、以下のことを明かにすることができた。 1.Prosタンパク質の局在と非対称分配にはProsタンパク質上のシグナルが必要である。 キイロショウジョウバエの卵細胞や初期胚では細胞の極性を決定する因子が細胞内に非対称に分布するが、これらの因子の局在はmRNA上のシグナルにより制御されることが知られている。pros遺伝子についてこの可能性を調べる目的で以下の実験を行った。 pros cDNAを核移行シグナル(NLS)を持ったLacZ遺伝子(NLSLacZ)にin-frameでつないだ融合遺伝子を作製し、神経芽細胞で発現させ抗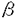 -galactosidase抗体により融合タンパク質の振舞を観察した。この融合タンパク質は内在性のProsタンパク質と全く同様に神経芽細胞で非対称に局在したのち、分裂に伴い神経母細胞へ分配された。 -galactosidase抗体により融合タンパク質の振舞を観察した。この融合タンパク質は内在性のProsタンパク質と全く同様に神経芽細胞で非対称に局在したのち、分裂に伴い神経母細胞へ分配された。 次に、LacZ遺伝子の後ろにpros cDNAをout-of-frameでつなぎ、-galactosidaseとpros全長の融合mRNAができるけれどもタンパク質は-galactosidaseまでしか翻訳されないようにデザインした融合遺伝子を神経芽細胞で発現させた。この場合には、-galactosidaseは神経芽細胞で均等に分布した後、分裂に伴い二つの娘細胞に等分配された。これらの結果は、-galactosidaseを非対称に局在させた後、分配するにはProsタンパク質そのものの上のシグナルが必要であり、胚極性の決定因子を細胞内で局在させる仕組とは異なることを示している。 2.非対称分配の活性を持つ領域の同定(’非対称’領域)。 -galactosidaseを非対称に分配する活性を持つ領域を系統的に探す目的で、次の実験を行なった。pros cDNAの翻訳領域を順次末端欠失したシリーズと、断片化したシリーズをNLSLacZ遺伝子5’側にin-frameでつないだ一連の融合遺伝子を作製し、これらの遺伝子産物の振舞いを調べてみた。 その結果、709番から943番の245アミノ酸(’非対称’領域)からなる領域に-galactosidaseを神経芽細胞内で非対称に局在させた後、神経母細胞に分配する活性があることが分かった。この’非対称’領域と-galactosidaseの融合タンパク質は、pros遺伝子の欠失変異株の胚でも非対称分配される。このことから、この融合タンパク質が野生胚で非対称に分配される際、内在性の完全なProsタンパク質が何らかの寄与をしている可能性を否定することができた。 3.Prosタンパク質の非対称分配に必須な配列の決定。 ’非対称’領域のどこに重要な情報が存在するかを調べる目的で以下の実験を行なった。 ’非対称’領域をC末から41アミノ酸削った領域( C1)と-galactosidaseの融合タンパク質は非対称分配される。ところが、このC1から更に32アミノ酸削った領域(C2)と-galactosidaseの融合タンパク質はもはや非対称分配されない。このことから、この32アミノ酸の中に非対称分配に必須な情報が含まれることがわかった。 C1)と-galactosidaseの融合タンパク質は非対称分配される。ところが、このC1から更に32アミノ酸削った領域(C2)と-galactosidaseの融合タンパク質はもはや非対称分配されない。このことから、この32アミノ酸の中に非対称分配に必須な情報が含まれることがわかった。 ’非対称’領域のC末端欠失変異の解析で同定された32アミノ酸以外に必須な領域がないかを検討するために以下の実験を行なった。ホメオドメイン以降を欠いたProsタンパク質(H領域;アミノ酸の1番から1237番)と-galactosidaseの融合タンパク質(HLacZ)は非対称分配される。このHの中の’非対称’領域に短い欠失を導入した一連の融合遺伝子を作製し、融合タンパク質の振舞を観察した。その結果、C末端欠失変異の解析で同定された必須な32アミノ酸の領域に、18および9アミノ酸の欠失を持つ融合タンパク質は非対称分配の活性を失うが、この領域以外の場所に短い欠失を持つ融合タンパク質は非対称分配の活性を保持していた。このことから、32アミノ酸からなる限定された領域がProsタンパク質の非対称分配に必須であることが判明した。 Homology searchの結果、この必須な配列に相同な配列が、Prosタンパク質と同様に非対称分配されるNumbタンパク質にも存在することが判明した。 4.pros遺伝子の発現は外胚葉由来の神経系だけでなく内胚葉由来の腸の原基でも認められるが、この組織の細胞が非対称分裂によって小さな細胞を生み出す過程で、Prosタンパク質が非対称分配されることがわかった。非対称分裂により生じた小さな細胞(adult midgut precursor)では、Prosタンパク質は核に局在している。 まとめと考察 細胞分裂を経て異なる二つの細胞が生まれるメカニズムとして、一方の娘細胞にのみintrinsic factorが非対称に分配される機構が想定され、実際に、protein kinaseや膜結合因子の分配が報告されている。prosperoは転写因子としては非対称に分配される初めての例である。これは、二つの娘細胞が異なる遺伝子発現をするために、転写因子が直接非対称に分配されるという新しいメカニズムである。 それでは、何故Prosタンパク質は非対称に分配されるのだろうか。prospero遺伝子が転写因子として機能する神経母細胞は、寿命の短い前駆細胞である。このような短寿命の細胞で一過性に活性を発揮するためには、de novoの産物の合成では間に合わないことが予想される。形成されたばかりの神経母細胞にProsタンパク質を供給するために、神経芽細胞で合成したProsタンパク質を輸送することは大変都合の良い仕組であると考えることができる。 本研究で、私は、 1.非対称な分布と、分配にはProsタンパク質上のシグナルが必要なこと、 2.そして非対称分配の活性を持つ領域を同定し、 3.その中に必須な機能を担う32アミノ酸の配列が存在すること、 を明らかにした。 この32アミノ酸からなる配列の中には、Prosタンパク質とNumbタンパク質に共通なモチーフが見つかることから、これら二つのタンパク質は共通の分子機構によって非対称に分配されることが示唆された。さらに、神経系以外でもProsタンパク質の非対称分配が認められることから、この仕組が発生の過程でかなり一般的に機能するものと考えられる。本研究で、同定した新しい機能ドメイン(非対称領域)とその中の必須なモチーフは、これまでにあまり知られていなかった細胞分裂に伴うタンパク質の非対称分配の分子機構の解明に、貴重な足掛りを与えるものである。 |