大腸がんは欧米型の食生活を行っている先進国で発生率の高い悪性腫瘍で、我国においても近年増加の傾向にあり、その発生予防に関する研究は重要な課題となっている。大腸発がん因子あるいは発生予防の研究にはラットあるいはマウスを用いた発がん実験が有効な方法としてあげられるが、著者は動物発がんモデルを応用して以下のような事実を明らかにしてきた。発がん物質のメチルニトロソウレアをラットに注腸し大腸癌を誘発する実験モデルを用い、ヒト等の腸内で産生される突然変異物質であるフェカペンテン12がプロモーター活性をもつことを明かにした。また、強力な抗酸化作用を有し、コレステロール低下薬として臨床で用いられているプロブコールがベンツピレンで誘発した大腸粘膜における前がん病変であるAberrant crypt foci(ACF)の形成を抑制することを見いだした。 一方、最近の発がんメカニズムの研究から、化学発がん剤投与や放射線照射等によるDNA傷害を細胞に与えた場合、細胞回転の制御機構やアポトーシスが、がん化の過程に重要な役割を果たしていることが明らかにされてきた。細胞の複製時に発がん剤に対する感受性やDNAの突然変異率は増大するが、組織内でDNA傷害を受け、転写不能に陥った細胞をアポトーシスという形で除去する生体防御機構が働いている。アポトーシスは壊死とは異なり、積極的に特定の細胞を除去するプロセスであり、核クロマチンの凝集から始まり、DNAの断裂へと進行しアガロース電気泳動では180bpのDNAフラグメントとして観察される。 p53がん抑制遺伝子はヒトの腫瘍中で最も高頻度に突然変異の認められる遺伝子で、そのコードする核タンパク質はアポトーシスの誘導や細胞回転の制御、DNA修復の進行をつかさどることが知られ、発がん過程を抑制する中心的な存在であると考えられている。本研究では、これまで主に培養細胞を用いて研究されてきたこれらの機構を、生体モデルを用いて解明することを目的とした。生体モデルとして用いた化学発がん剤投与を受けたマウスの大腸粘膜における細胞回転、アポトーシスの誘導及びp53タンパクの生体内での発現を解析し、また同時に、マウス大腸発がんのイニシエーション過程でのp53タンパクの役割についてもノックアウトマウスを用いて検討を行った。 生体モデルとしては大腸腫瘍を特異的に誘発することの知られるアルキル化発がん剤であるアゾキシメタン(AOM,20mg/kg)を腹腔内投与したC57BL/6マウスを用いた。アポトーシスの検討は投与後マウスを経時的に屠殺し、大腸よりDNAを抽出し、アガロース電気泳動を行いDNAのフラグメンテーションを観察した。組織は10%ホルマリン液固定後にパラフィン切片を作製し、各種の免疫組織化学的手法を含む病理組織学的検討を行った。アポトーシスはDNAフラグメンテーションとして投与後3時間から観察され始め、48時間後まで観察可能であった。また、AOMの投与量を2.5mg/kgから20mg/kgまで変化させた場合、AOM 5mg/kg投与からアポトーシスは認められ、発がん剤の投与量に依存して増強し、また同様のサンプルに対し、ヘマトキシリンーエオジン染色とアポトーシスの特異的染色法であるTUNEL法を用いた染色を行ったところ、大腸1陰窩あたりのアポトーシスに陥った細胞数はAOM投与後6時間で最大となり、その後減少した。同時にマウスp53タンパクに対するポリクローナル抗体を用いた免疫染色を行ったところ、p53タンパクが核へ蓄積した細胞数の経時的変化とアポトーシス細胞の出現数の増減はよく一致した。さらに、アポトーシスに陥った細胞とp53抗体陽性の細胞は共に、大腸陰窩基底部の増殖帯の部位に一致して認められた。大腸粘膜における化学発がん剤AOM投与後の細胞増殖の変化については抗BrdU抗体を用いた免疫組織染色で検討した。その結果、AOM投与後12時間目で有意なS期細胞数の抑制が観察された。これは培養細胞を種々のDNA傷害物質で処理した時、細胞回転が複製前のG1期に停止する所見と一致し、細胞のDNA傷害を修復するための合目的性のある反応とみなされる。AOM投与後のBrdU染色陽性の細胞数抑制は48時間目に解除され、S期細胞が観察された。この結果はすでに報告されているマウスに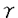 線の全身照射を行い得られた大腸粘膜の変化についての報告と類似していた。 線の全身照射を行い得られた大腸粘膜の変化についての報告と類似していた。 以上の実験により確認された生体内におけるp53タンパクによる細胞回転の制御、アポトーシスの誘導に加え、さらにがん化への関連を明確にする目的でp53遺伝子ノックアウトマウスを用い、AOM投与による影響を観察した。AOM(20mg/kg)投与6時間後の大腸陰窩上皮細胞の1陰窩あたりのアポトーシス数は、p53(-/-)マウスでは0.3±0.04個とほとんど認められなかった。それに対し正常p53(+/+)マウスでは5.1±0.8個、ヘテロ接合体p53(+/-)マウスでは2.5±0.3個とp53タンパク量に依存した値を示した。また、BrdU標識S期細胞の検討を行ったが、6時間後ではAOM投与によりp53(+/+)マウス及びp53(-/-)マウスの両者で類似したS期細胞数の低下が認められ、p53蛋白とは無関係に起こった現象と考えられた。しかし、AOM投与48時間後に上昇したS期細胞数はp53(-/-)マウスではp53(+/+)マウスに比べ有意に高く、これらの反応の制御にp53蛋白が関係していることが窺われる結果が得られた。 上述した如く、p53(-/-)マウスの大腸粘膜において、AOM投与によるアポトーシスの抑制と投与後48時間でのDNA合成の亢進が観察されたことから、更に大腸粘膜における前がん病変であるACF数及びBrdU陽性細胞について検討を行った。p53(-/-),(+/-),(+/+)マウスに各々AOM 7mg/kgを腹腔内に週1回4週間投与し、最終投与から1週間後にマウスを屠殺し、大腸粘膜のACF数を算定した。(+/+)マウス及び(+/-)マウスの大腸粘膜に発生したACF数はほぼ同程度(6.4±4.4、7.4±4.1)であったが、(-/-)マウスでのACF数は少数(0.8±1.0)であり発生が抑制される結果となった。この結果はp53(-/-)マウスを用いたジメチルベンツアントラセン誘発皮膚腫瘍でも腫瘍発生が(-/-)マウスでは(+/+)または(+/-)マウスに比べ低いという報告と類似した結果となった。また、この時の大腸粘膜のBrdU陽性細胞数は(-/-)マウスで正常マウスに比べ有意に高かった。本研究では生体内で化学発癌物質を投与した動物の大腸についてアポトーシス、p53タンパクの発現、S期細胞数の変化を検出、解析し以下の点を明らかにし得た。 1.アポトーシス出現、p53タンパクの発現の経時的変化は、大腸粘膜では基本的には培養細胞の研究から得られた結果と一致していたが、大腸陰窩の増殖帯にのみ観察されることが明かとなった。このことより生体内では常時回転していない細胞はp53依存性のアポトーシスに陥いらないことが推測された。 2.発癌物質AOMをp53遺伝子ノックアウトマウスに投与して、アポトーシスの出現を正常マウスと比較したところノックアウトマウスでは大腸粘膜にはアポトーシスは生じず、AOM投与48時間後に観察されるS期細胞数の増加率は正常マウスに比べノックアウトマウスでは有意に高かった。 3.発癌物質AOMをp53遺伝子ノックアウトマウスに連続投与し、大腸粘膜の前がん病変ACF数を算定したところ、ノックアウトマウスではその発生は正常マウスに比べ抑制されたが、S期細胞の増加が観察された。 |