Tリンパ球に対して抗原提示を行うことが知られている単球やBリンパ球は、刺激によりスーパーオキシドアニオン(O2-)を産生する。すなわち前者では細菌貪食時や可溶性の刺激物存在下で、また後者では表面抗原を架橋したり可溶性の刺激物の存在下でO2-産生系が活性化される。これに対して抗原提示性細胞として知られている樹状細胞のO2-産生系については良く知られていない。ヒト単球をIL-4とGM-CSFの存在下で培養すると樹状細胞様細胞へ分化する。このようにして分化した樹状細胞様細胞のO2-産生能を調べたところ、この細胞は既にO2-産生能を失っていることを見い出した。そこで、その分化過程におけるO2-産生能の変化を解析することにした。 ヒト末梢血より分離した単球はGM-CSFとIL-4の存在下で1週間培養するとほとんどの細胞は非附着になり、樹状細胞の特徴を示す。これらの細胞は非特異エステラーゼ陰性で、非貪食性である。CD1,CD11c,MHCIIの発現が高く、CD14とCD71の発現は抑えられており、T細胞に対して高いMLR(Mixed Lymphocyte Reaction)刺激活性を示した。一方、GM-CSFのみの存在下で培養すると単球はマクロファージ様細胞に分化し、またサイトカインなしまたはIL-4のみを加えた場合に単球は分化せず、ほとんどが死滅する。 このように分化した樹状細胞様細胞についてPMA(Phorbol myristate acetate)刺激によるO2-産生能をlucigenin依存的なケミルミネッセンス法で調べたところ、その活性を認めなかった。これに対して、培養前の単球は細胞数依存的に高いO2-産生能を示すことが確認された。そこで、シトクロムc還元法でこのO2-産生能の変化を時間を追って追跡した(図1)。GM-CSFとIL-4の存在下で2日間培養した細胞はO2-産生能が低下し、7日後に得た樹状細胞様細胞は完全にO2-産生活性を失っていた。これに対してGM-CSFのみの存在下で培養した単球はマクロファージ様細胞に分化したがそのO2-産生能に変化は見られなかった。 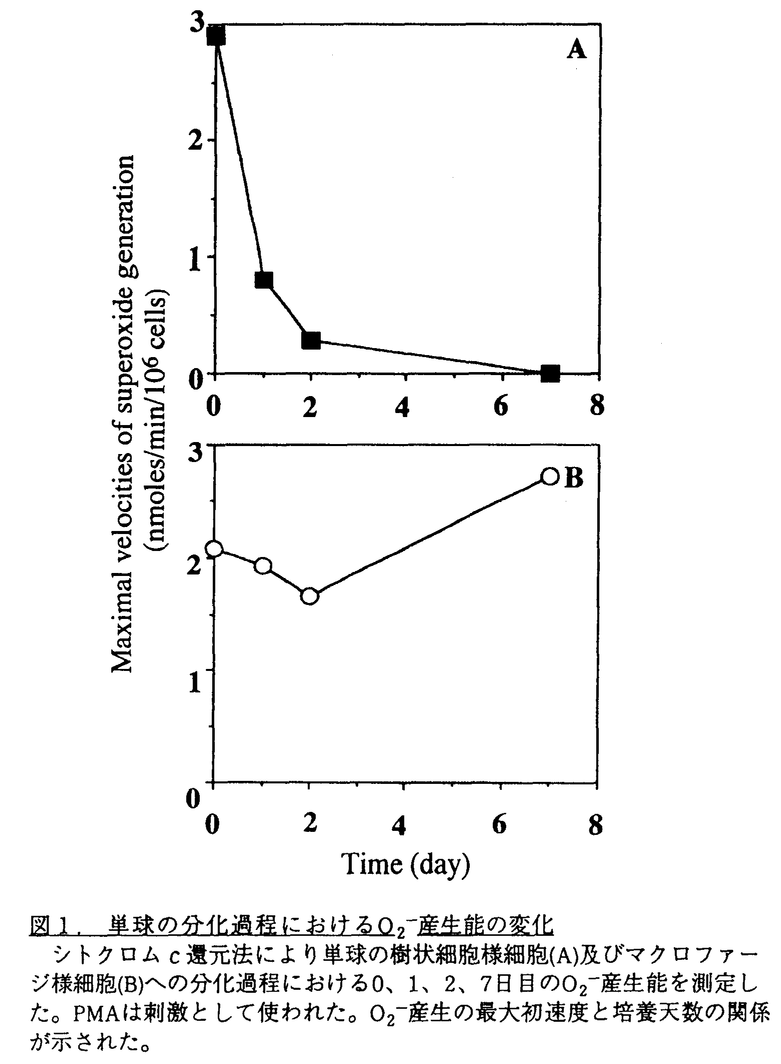 図1.単球の分化過程におけるO2-産生能の変化シトクロムc還元法により単球の樹状細胞様細胞(A)及びマクロファージ様細胞(B)への分化過程における0、1、2、7日目のO2-産生能を測定した。PMAは刺激として使われた。O2-産生の最大初速度と培養天数の関係が示された。 図1.単球の分化過程におけるO2-産生能の変化シトクロムc還元法により単球の樹状細胞様細胞(A)及びマクロファージ様細胞(B)への分化過程における0、1、2、7日目のO2-産生能を測定した。PMAは刺激として使われた。O2-産生の最大初速度と培養天数の関係が示された。 単球の樹状細胞様細胞への分化過程におけるO2-産生能低下についての機構を明らかにするためにO2-産生系構成蛋白質に対する抗ペプチド抗体を用いてそれぞれの蛋白質の量の変化を免疫ブロッテング法で解析した。O2-産生には形質膜の特殊なシトクロムを形成する二つのサブユニット91-kDa(gp91)及び22-kDa(p22)の蛋白質と細胞質性の47-kDa(p47)、65-kDa(p65)、Rac-p21各蛋白質が必要である。4回の実験結果をBioImage Analyzerで定量し、図2にその経時的変化を示したが、gp91とp22の量は培養2日目まで急激に減少した。これに対して細胞質性のp47とp65の量は培養中一定して変わらなかった。一方、Rac-p21は樹状細胞様細胞には単球と同じレベルで存在した。さらにシトクロムを細胞外から認識する単クローン抗体7D5を用い、フローサイトメトリー法で単球と樹状細胞様細胞のシトクロムの変化を調べたところ、免疫ブロッテングと一致して、樹状細胞様細胞において明らかな減少が認められた。免疫ブロッテングに用いた抗体はシトクロムの各サブユニットのC末端を識別するので単球から樹状細胞様細胞への分化に際してはこの部分がまず分解していくものと考えられる。 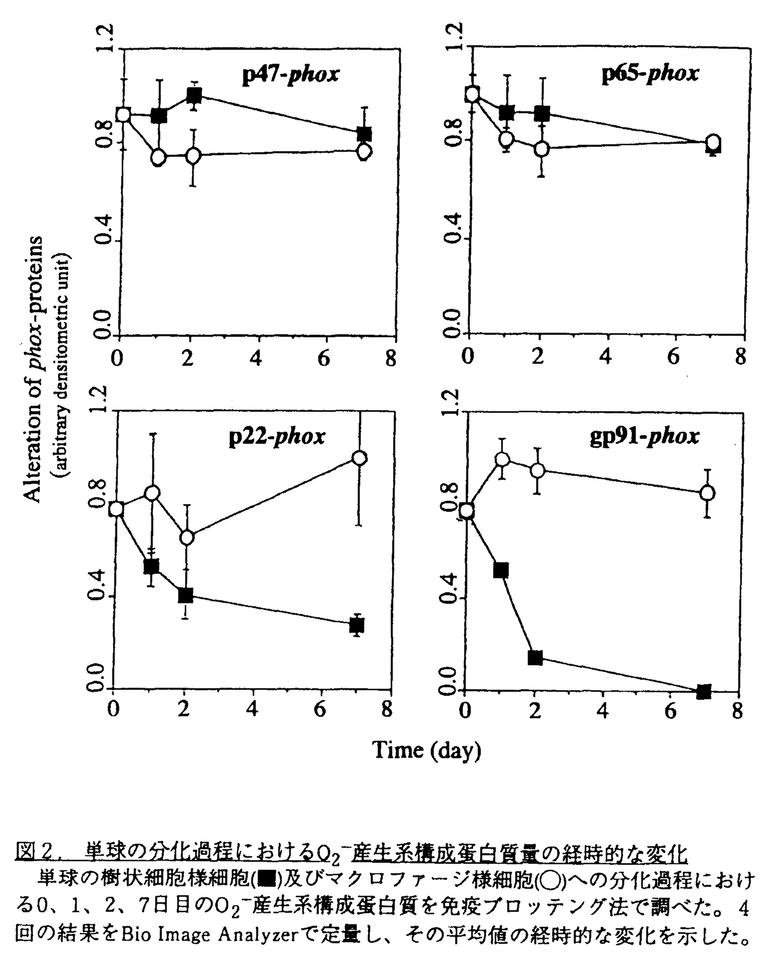 図2.単球の分化過程におけるO2-産生系構成蛋白質量の経時的な変化単球の樹状細胞様細胞(■)及びマクロファージ様細胞(○)への分化過程における0、1、2、7日目のO2-産生系構成蛋白質を免疫ブロッテング法で調べた。4回の結果をBio Image Analyzerで定量し、その平均値の経時的な変化を示した。 図2.単球の分化過程におけるO2-産生系構成蛋白質量の経時的な変化単球の樹状細胞様細胞(■)及びマクロファージ様細胞(○)への分化過程における0、1、2、7日目のO2-産生系構成蛋白質を免疫ブロッテング法で調べた。4回の結果をBio Image Analyzerで定量し、その平均値の経時的な変化を示した。 さて、このように一旦分化し浮遊細胞となった樹状細胞様細胞はIL-4非存在下で培養を続けると再度付着細胞となりマクロファージ様細胞の形態を示すことが明らかとなった。そこで、マクロファージのマーカーである非特異エステラーゼ活性及びNBT(Nitroblue tetrazolium)還元法でそのO2-産生能を調べた。樹状細胞様細胞は両者とも陰性であったが、再び付着するようになった細胞は非特異エステラーゼ活性は陽性で、MBT還元活性は陰性であった。この結果は樹状細胞様細胞は条件により再度マクロファージ様細胞に(脱)分化するが、O2-産生活性は回復できないことを示している。 好中球と同様に、単球やマクロファージはその細胞表層にユニークなO2-産生系を持ち、刺激を受けるとO2-を産生する。最近、これらと同じO2-産生系がBリンパ球にも存在することが当研究室から報告された。末梢血や扁桃におけるBリンパ球のO2-産生能はsIgMとsIgDの様な表面抗原の架橋により活性化されることから、そのO2-産生と抗原提示性と結び付く可能性が推測された。抗原提示性細胞として知られる樹状細胞は各組織に広く分布する。本研究で示したように単球から分化した樹状細胞様細胞にはO2-産生活性は認められず、また組織に存在する樹状細胞にはシトクロムが検出されなかったので、この仮説は少なくとも樹状細胞には適用されない。 単球またはマクロファージをサイトカイン非存在下で培養するとO2-産生能が抑さえられることが報告されているが、そのような条件下ではこれらの細胞はアポトーシスを起こすことが明らかになっている。一方、単球やマクロファージにIL-4を与えるとO2-産生能が低下すること、さらに最近豚の肺胞マクロファージではそれがgp91のmRNAの発現低下によることが示された。IL-4の存在下で単球やマクロファージはアポトーシスをおこすためO2-産生能の低下はアポトーシスの結果と考えられたが、今回の結果はO2-産生活性の低下とアポトーシスが関係がないことを示している。 今回示したように単球やマクロファージ様細胞は樹状細胞様細胞に分化するが、IL-4非存在下ではマクロファージ様細胞に戻る。このことは生体内でも単球やマクロファージはサイトカイン(IL-4など)の存在下で樹状細胞様細胞になったりマクロファージ様細胞になったりする可能性を示している。しかし、このように樹状細胞様細胞由来のマクロファージ様細胞は元の単球やマクロファージと違って、O2-産生は出来ないため異なった役割を果たす可能性もある。 |