TGF-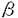 RII遺伝子の発現解析
RII遺伝子の発現解析 TGF-RII遺伝子の発現と細胞の形質転換の関連を検討した。各種培養細胞から全RNAを抽出し、TGF-RIIのcDNAをプローブとして、ノーザン法を行った。TGF-RIImRNAの発現は胃癌(MKN28)、子宮頚癌(HeLa)、前骨髄球性白血病(HL60)の各細胞株では非常に低く、肝細胞癌細胞株(HepG2)では非常に多くなっていた。
DNA型癌ウイルスのアデノウイルスやSV40で形質転換したヒト線維芽細胞株(293、TIG-wt)でも、形質転換前のTIG細胞よりTGF-RIImRNAの発現が1/7以下になっていることを見出した。
さらに、SV40をCV1細胞に感染(m.o.i.10)させると、TGF-RIImRNAのレベルは感染後24時間では変化しないが、感染後48時間では著名に減少していることを見出した。SV40の感染に伴うTGF-そのもののmRNAの発現変化は見られなかった。
上記の結果から、多くの癌細胞でTGF-RIImRNAの発現が低いことを確認した。さらに、SV40による形質転換後のTIG細胞のTGF-RIImRNAの発現の減少も見い出した。また、SV40感染後のCV1細胞におけるTGF-RIImRNAの発現も減少するが、この発現減少は比較的早い時期に現われたことで、この変化は染色体上の変異によるものではないと考えられた。同時に、SV40感染によりTGF-mRNAの発現変化は見られないことから、変化はmRNAのレベルが全体として下がるせいではなくTGF-RIIに特異的と考えられる。TGF-RIIはTGF-より細胞の形質転換に深く関係している可能性が高い。HepG2細胞におけるTGF-RIIの高い発現に関してはその機構は不明である。
TGF-RIIプロモーターのクローニングと解析 TGF-RIImRNA発現調節機構を調べるために、この遺伝子のプロモーター領域の分離を行った。ファージSharon4Aのヒトゲノムライブラリーから、TGF-RIIcDNA5’-末側の337bp断片をプローブとしてプラークハイプリダイゼーションを行い、約30万クローンをスクリニーグし、陽性クローンを得た。その中のゲノム断片の制限酵素マップをサザン法で明らかにし、TGF-RIIcDNAの5’-末開始点上流1.1k、下流330bpの1430bpの領域を含んでいることを確認した。
プロモーター活性を測定するルシフェラーゼアッセイ用plasmid pGL2Basicに1430bpの領域をつなぎ(pGL32)、ルシフェラーゼアッセイにより、このクローンはプロモーター活性があることを見い出した。
次に、pGL32の塩基配列をALFシークンサー(Pharmcia)を使い、ジデオキシ法で決定した。塩基配列の特徴はTATAボックス及びCAATボックスが存在しない。また、転写因子結合エレメントとしては、Sp1配列がcDNAの第一塩基を+1として、-25→-20と-142→-137の2カ所にある。また、-185→-180にAP1配列が存在する。
プロモーター活性に必要な最小領域を決定するために、pGL32から種々の欠失株、変異株を作製し、CV1細胞でルシフェラーゼアッセイを行い、各欠失、変異株の転写活性を測定した。その結果、転写開始領域付近にそれぞれ単独でプロモーターとして機能しうる約80bp程度の2つの領域が互いに接していることがわかった。これらのプロモーター活性がある領域につき、TGF-RII遺伝子のcDNAの第一塩基を含むものをP1、その上流に存在するものをP2と命名した。
まず、P1の領域について解析した。-22→+55と-15→+330領域にはプロモーター活性があったが、-48→+17と-9→+340領域は活性がなかった。これはP1の活性には+18→+55及び-22→-9の領域中に不可欠な塩基配列が存在することを示している。また-22→+35の領域のプロモーター活性は-22→+55の領域の活性と比べ1/3以下に低下した。これは、P1のプロモーター活性機能を担う塩基配列の末端がが下流側の+35→+55の領域に存在することを示している。
以上をまとめると、P1のプロモーター活性を担う領域は-15から+55までの領域中にあり、その中でも-15→+35の領域がこのプロモーター活性が不可欠であることを明らかにした。
次にP2の領域について解析した。-137→-22領域は-1.1k→-22領域と変わらぬプロモーター活性を示した。-105→-22領域は-137→-22領域の1/2プロモーター活性があった。-105→-48及び-1.1k→-48の領域の活性は-105→-22領域のと比べ1/3しかなかった。-48→+17領域はプロモーター活性がなっかた。これらの結果は、P2は-137→-22の領域にある。その中に-137→-105と-48→-22の領域はプロモーターの活性に影響を持つ部分である。
まとめ、TGF-RII遺伝子はP1ならびにP2の2つのプロモーターによって発現を制御されていると言える。P1のプロモーター活性がある領域には、これまで報告されている特異的塩基配列は転写因子モチーフが存在しない。従って、今回見出したTGF-RIIプロモーターは、まだ知られていない構造を持つ可能性がある。
 II型受容体プロモーターの分離及び機能領域の解析
II型受容体プロモーターの分離及び機能領域の解析