HLA(ヒト白血球抗原)分子はほとんどすべての細胞表面に発現されている多型性に富む糖タンパク質であり、免疫反応の制御や自己と非自己の識別において重要な役割を演じている。 HLA遺伝子群は第6染色体の短腕上のp21.3バンドにある、約4,000,000塩基対からなるHLA複合領域と呼ばれる領域に存在する。この領域には多数の発現、非発現遺伝子があり、クラスI、クラスII、クラスIII領域に分けられる。クラスI領域にはHLA-A,B,Cをコードしている遺伝子が、またクラスII領域にはHLA-DR,DQ,DPをコードしている遺伝子が含まれる。 HLA遺伝子群は極めて高い多型性を示す。それぞれの遺伝子座には多数の対立遺伝子があり、現在までHLA-A,B,C,DR,DQ,DPの対立遺伝子としてそれぞれ61,128,36,152,25,66種類が明らかにされている。さらにHLA遺伝子の興味深い点はこれらの対立遺伝子間の差異の大部分が非同義置換であることである。したがってこれらの対立遺伝子の産物のほとんどは異なったアミノ酸配列を持つ。 HLA抗原は生体に発現されている抗原の中で最も強い抗原性を示し、HLA抗原の適合、不適合が輸血や移植において臨床結果を大きく左右する問題となる。 輸血においては頻回の輸血により患者に抗HLA抗体が産生され、HLA抗原を発現している血小板の輸血に対し不応状態となる。またドナーと患者のHLA不一致により起こる輸血後移植片対宿主病(PT-GVHD)は極めて致死率の高い重篤な輸血副作用である。 HLA抗原の異なるドナー-患者間の移植では強い拒絶反応やGVHDを引き起こす。HLA抗原のタイピングはこれまで血清学的方法で行われてきたが、血清学的には区別できない抗原があり、移植において拒絶反応やGVHDの原因となりうる。血清学的には完全に適合した骨髄の移植において拒絶反応が起きた症例がアメリカにおいて報告されており、この症例ではドナーのHLA-B44はB*4403、患者はB*4402であり、B*4403特異的細胞傷害性T細胞が産生されていた。このような知見から対立遺伝子のタイピングの必要性が認識されるに至った。 HLA抗原はそのX-線解析により立体構造が解明され、クラスI抗原では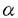 1と2ドメイン、クラスII抗原では1と 1と2ドメイン、クラスII抗原では1と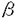 1ドメインにより形成される溝に自己または非自己のペプチドを結合することが明らがとなっている。このHLA抗原とペプチドの複合体がT細胞、NK細胞に認識され免疫応答が開始される。HLA抗原の多型性はペプチドを結合する溝を構成しているドメインに集中している。 1ドメインにより形成される溝に自己または非自己のペプチドを結合することが明らがとなっている。このHLA抗原とペプチドの複合体がT細胞、NK細胞に認識され免疫応答が開始される。HLA抗原の多型性はペプチドを結合する溝を構成しているドメインに集中している。 したがってHLA抗原の異なる個体ではT細胞、NK細胞レセプターの特異性が異なり、特定の抗原に対する免疫反応が異なる可能性が考えられる。このようなHLA抗原の特徴から対立遺伝子と疾患との関連が研究されている。例えばHLA-B27は強直性脊椎炎の患者に陽性者が高いことが知られており、その発症の機構を解明するため詳細な研究がなされている。これまでの研究では、B27をコードする対立遺伝子としてこれまで9種類(B*2701-B*2709)が確認されているが、B*2703,B*2706を持つ個体ではこの疾患に対して感受性が低く、B*2704,B*2705では感受性が高いことが報告されている。 HLA抗原はその高度な多型性のため、人類学の分野においても有用な知見を提供する。これまでいくつかのHLA抗原対立遺伝子については民族集団によりその頻度が異なることが明らかとなっている。さらに各民族集団における対立遺伝子分布を決定することにより、民族間の遺伝的な関係の推定が可能になると考えられる。 以上のようにHLA抗原系を詳細に解明することは輸血、移植などの臨床医学、自己免疫疾患の発症機構などの基礎医学あるいは人類学、遺伝学などの分野にも有用な情報を提供する。このような見地から、クラスII抗原については遺伝子解析が比較的用意であることもあり、ほとんどの対立遺伝子が決定されてきた。しかし、クラスI遺伝子については、その多型性を示す領域がクラスII遺伝子に比べて長く、特にHLA-B遺伝子はクラスI遺伝子の中でも特に多型性が高いことから、未解析の対立遺伝子が残されている。またこれまでの対立遺伝子の解析はそのほとんどがCaucasianのものであり、アジア系集団についての報告は極めて少ない。 そこで本研究では、アジア系集団のHLA-B遺伝子座、特にHLA-B40およびB15グループの多型性、分子構造を調べるため、16種類のHLA-B対立遺伝子の塩基配列を決定した。このうち4種類(B*4007,B*1528,B*1529,B*1802)はこれまでに報告のない新しい対立遺伝子であった。 さらに対立遺伝子の各民族集団における分布を調べるため、B40-,B15-,B13-,B18-陽性個体の試料、それぞれ284,131,57,18検体を得て、PCR-SSO法およびPCR-SSCP法を用いて対立遺伝子を同定し、つぎの結果を得た。 (1)HLA-B40グループ(B60,B61)は日本人、北方漢民族、満族、韓国人、中国に在住の朝鮮民族(以下朝鮮族)、ブリヤート(シベリヤに住むモンゴル民族)およびモンゴル人について調べた。B60抗原はB*40012に、B61抗原はB*4002,B*4006および頻度は低いがB*4003にコードされていた。これらのB61対立遺伝子頻度には民族差が見られた。ブリヤート、モンゴル人のB61ではB*4002が高頻度であったが、満族と朝鮮民族ではB*4002とB*4006がほぼ同頻度であり、日本人と韓国人ではB*4006に比べB*4002が高頻度に見られた。B*4003は日本人、朝鮮民族およびモンゴル人に見出されたが、韓国人、ブリヤート、満族,北方漢民族には見出されなかった。本研究で見出されたB40グループの新しい対立遺伝子であるB*4007は日本人でのみ見出された。 (2)日本人とアジア人(タイ、ベトナム、インドネシア、マレーシア)のB15グループ(B62,B70,B75,B77)を調べたところ、8種類のB15対立遺伝子が検出された。B62をコードしているB*1501はアジア人に頻度の高い対立遺伝子であった。B15NをコードしているB*1502は中国人では比較的高頻度であったが、日本人、韓国人、朝鮮民族ではまれであった。B75VとB70をそれぞれコードしているB*1511とB*1518も見出された。オーストラリアアボリジニで報告さていたB*1521もアジア人から見っかった。B77をコードしているB*1513はタイ人にはよく見られる対立遺伝子であったが、日本人、韓国人、朝鮮民族ではまれであった。本研究で始めて見出された対立遺伝子であるB*1528とB*1529は3人の日本人と民族不明の1人で見つかった。 (3)2つのB13スプリット抗原が日本人およびタイ人から報告されていた。塩基配列決定により、この内1つはB*1301に、もう1つはB*1302にコードされていることが明らかとなった。これまでの報告ではB*1302はアジア人には存在しないとされていたが、今回の研究でアジア人での存在が確認された。 (4)日本人、中国人、ブリヤート、韓国人、朝鮮民族および東南アジア人を調べたところ、新しい対立遺伝子であるB*1802がタイ人、インドネシア人、ベトナム人からそれぞれ1検体づつ、3検体見つかった。 (5)本研究で確認されたほとんどの対立遺伝子は他のHLA遺伝子座抗原と強い関連を示した。B*4002とB*4003はCw10と強い連鎖不平衡を示し、B*4006はCw*0801と強く関連した。B*1301はA2,Cw10,DR12,DQ7と、B*1302はA30,Cw6,DR7,DQ2と強く関連した。B*1502はCw8と強く関連したが、B*1521はA34,Cw6と強く関連した。アジア人で見つかった3検体のB*1802陽性検体はすべてHLA-Cw7,DR12,DQ7を持っていた。 以上の研究によりHLA-B遺伝子座、特に日本人およびアジア人のB40とB15グループの塩基配列およびアミノ酸一次構造、さらに抗原と対立遺伝子の対応が明らかとなった。幾つかの対立遺伝子は民族集団により異なった分布を示しており、ほとんどの対立遺伝子が他のHLA遺伝子座抗原と強い関連を示した。 骨髄移植におけるHLAのタイピングはクラスIIについては対立遺伝子レベルでのタイピングが可能となっている。その結果、ドナー-患者間のHLAが血清学レベルで一致していた場合に比べ、対立遺伝子レベルで一致していた場合の方が臨床成績が明らかによいことが報告されている。また輸血においてGVHDは最も重篤な副作用である。GVHD発症の機構を明らかにするためにはやはり対立遺伝子レベルでの解析が必要である。現在、クラスIのタイピングは血清学的手法に依存しているが、今回得られたクラスI抗原と対立遺伝子の関係およびそのタイピング法の確立はクラスIのDNAタイピングを可能とし、移植の臨床成績および輸血の安全性向上に必須の情報を提供すると考えられる。 HLAの多型性とその機能、疾患発症との関係に関する研究においてもクラスIIの対立遺伝子レベルでの解析が行えるようになったことにより、関連がより明確になり、詳細な研究が進められている。本研究で得られたクラスI領域の知見はこのような研究分野にも役立つものと考えられる。 |