本研究は、Ca2+代謝において重要な役割を演じる破骨細胞のCa2+受容体(Ca2+sensor)の機能に関与すると考えられている細胞膜電位の制御機構を明らかにするため、ウサギ破骨細胞の単離培養系において、電気生理学的手法を用いて膜電位に関わるイオンチャネルの同定およびその制御機構の解析を試みたものであり、下記の結果を得ている。 1.これまでウサギ破骨細胞において報告されていた「細胞外Ca2+濃度の上昇により抑制される内向き整流K+チャネル」だけでは細胞外Ca2+濃度の上昇による膜電位の変化(約-80mVの静止膜電位から脱分極して約-20mVに固定される)を説明できなかったが、K+チャネルを抑制した条件で解析することにより、「細胞外Ca2+濃度の上昇により可逆的に活性化されるDIDS感受性外向き整流Cl-チャネル」の存在を明らかにした。従って、ウサギ破骨細胞の膜電位は、ほぼ生理的な細胞外Ca2+濃度下(1mM)ではK+の平衡電位に近い約-80mVにあるが、細胞外Ca2+濃度上昇により内向き整流K+チャネルが抑制されDIDS感受性外向き整流Cl-チャネルが活性化されることによりCl-の平衡電位に近い約-20mVに脱分極することが示された。 2.Ca2+と同じく細胞内Ca2+濃度を上昇させるCd2+やNi2+等の2価イオンと外向き整流Cl-チャネルの活性化について検討することにより、Ca2+以外の2価イオンがCa2+と同様に外向き整流Cl-チャネルを活性化すること、及び活性化の時間経過のパターンが細胞内Ca2+濃度の変化と類似していることを明らかにし、このCl-チャネルが細胞内Ca2+濃度の上昇を介して活性化される可能性を考えた。細胞内Ca2+を20mMのEGTAでキレートすると、細胞外2価イオンによるCl-チャネルの活性化は抑制された。以上より、細胞外Ca2+により誘発される外向き整流Cl-チャネルはCa2+-activated Cl-チャネルであることが明らかになった。 3.細胞内にGTP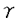 Sを投与したところ、細胞外Ca2+濃度が1mMでも、GTPSは外向き整流Cl-電流を誘発し、外向き電流の大きさが一定に達した後、細胞外Ca2+濃度を増加させてもさらなる電流量の増大は認められず、ここでDIDSを投与するとこの電流は抑制された。パッチ電極のEGTA濃度を20mMにして細胞内Ca2+をキレートしてやるとGTPSは外向き整流電流を誘発しなかった。以上の結果より、GTPS誘発電流は細胞外Ca2+誘発Cl-電流と同一であると結論され、よって細胞外Ca2+濃度上昇によるCl-チャネルの活性化にはG蛋白が関与していること、また、細胞内制御機構においてG蛋白の活性化のdownstreamに細胞内Ca2+濃度上昇が位置している可能性が示唆された。なお、百日咳毒素処理は細胞外Ca2+濃度上昇によるCl-チャネルの活性化を抑制せず、関与するG蛋白は内向き整流K+チャネルの抑制の場合と同様に百日咳毒素非感受性G蛋白と考えられた。 Sを投与したところ、細胞外Ca2+濃度が1mMでも、GTPSは外向き整流Cl-電流を誘発し、外向き電流の大きさが一定に達した後、細胞外Ca2+濃度を増加させてもさらなる電流量の増大は認められず、ここでDIDSを投与するとこの電流は抑制された。パッチ電極のEGTA濃度を20mMにして細胞内Ca2+をキレートしてやるとGTPSは外向き整流電流を誘発しなかった。以上の結果より、GTPS誘発電流は細胞外Ca2+誘発Cl-電流と同一であると結論され、よって細胞外Ca2+濃度上昇によるCl-チャネルの活性化にはG蛋白が関与していること、また、細胞内制御機構においてG蛋白の活性化のdownstreamに細胞内Ca2+濃度上昇が位置している可能性が示唆された。なお、百日咳毒素処理は細胞外Ca2+濃度上昇によるCl-チャネルの活性化を抑制せず、関与するG蛋白は内向き整流K+チャネルの抑制の場合と同様に百日咳毒素非感受性G蛋白と考えられた。 4.破骨細胞をokadaic acidで数分間前処置すると、細胞外Ca2+濃度上昇によりCl-電流が非可逆的に活性化された。また細胞外にCd2+を投与すると、非処置時には一過性であるCl-電流の活性化が持続性に認められた。以上より、このCa2+-activated Cl-チャネルの活性化にはリン酸化・脱リン酸化の過程も関与している可能性が示唆された。 以上、本論文は細胞外Ca2+濃度上昇がウサギ破骨細胞の膜のCa2+受容体を介して百日咳毒素非感受性G蛋白を活性化し、その結果内向き整流K+チャネルは抑制され、一方細胞内Ca2+濃度が上昇してCa2+-activated Cl-チャネルが活性化されることによって膜が脱分極するという一連の制御機構がある可能性を示した。この膜電位の変化はCa2+受容体の機能を変化させ、また細胞外からのCa2+流入を変えることにより細胞内Ca2+濃度を変化させている可能性がある。従ってCa2+受容体によるイオンチャネルの制御は破骨細胞の機能に重要な役割を担っている可能性があり、その生理機構の解明に重要な貢献をなすと考えられる本研究は学位の授与に値すると思われる。 | 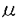 Mであった。
Mであった。