膵は内胚葉性上皮由来の臓器であり、十二指腸の背側と腹側に形成される各々1個の膵原基より発生し、胎生期には腹側膵原基が背方に移動して背側原基と融合して膵臓が形成される。完成された膵臓は、外分泌腺細胞からなる膵腺房、内分泌腺細胞からなる膵島(ランゲルハンス島)、導管系上皮細胞からなる膵管で構成される。そしてその細胞レベルの分化は、膵原基に存在する膵導管系の前駆細胞から膵外分泌腺細胞と膵内分泌腺細胞が分化するとされており、この膵の分化も他の臓器と同じように複数の増殖分化因子の協調作用により司られていると考えられている。1973年にLevineらによる胎児の移植片を使った実験で、膵分化を刺激する胎生期の分化因子の存在が示されたがその因子は未だ同定されていない。しかし、既知のペプチドで膵の分化を刺激するものは知られている。例えばTGF-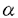 を膵に過剰発現させたトランスジェニックマウスでは膵導管の過形成が生じ、さらにアミラーゼを発現する膵外分泌腺様細胞への化生性変化が膵管細胞に生じる。また、TGF-とガストリンを同時に膵に発現させるとランゲルハンス島の細胞数が増加し、インスリンの発現量も増加することが示されている。また、実験的に作られたマウスの糖尿病の進展を抑えることが知られているニコチンアミドが、ヒトの胎児膵のB細胞の分化を促すことも最近示された。 を膵に過剰発現させたトランスジェニックマウスでは膵導管の過形成が生じ、さらにアミラーゼを発現する膵外分泌腺様細胞への化生性変化が膵管細胞に生じる。また、TGF-とガストリンを同時に膵に発現させるとランゲルハンス島の細胞数が増加し、インスリンの発現量も増加することが示されている。また、実験的に作られたマウスの糖尿病の進展を抑えることが知られているニコチンアミドが、ヒトの胎児膵のB細胞の分化を促すことも最近示された。 アクチビンAは、卵胞液中より単離精製された分子量25kDの蛋白質であり、その構造上の特徴によりTGF-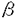 スーパーファミリーに属しており、同じTGF-スーパーファミリーに属するインヒビンのAサブユニットで構成される二量体構造をもっている。アクチビンは性腺以外にも、下垂体、神経系、骨髄、副腎、肝臓、膵などの様々な臓器での発現が知られており、局所において産生されオートクリン、パラクリン因子として細胞の分化や増殖および発生に大きな影響を及ぼしている。たとえばその作用として、赤芽球及び巨核球の分化誘導作用、神経栄養因子としての作用、またリンパ球、肝細胞、血管平滑筋細胞などの増殖制御因子としての作用が知られている。また発生段階では中胚葉の分化誘導因子としても働いている。このように、アクチビンAは発生の段階から分化・増殖をはじめとして様々な細胞調節機構に局所因子として関与する多機能性タンパク質である。そしてアクチビンAは、ラット膵においてラ氏島に特異的に発現しており、また胎生期の膵内分泌腺前駆細胞にも発現していることから、膵の分化および増殖において何らかの役割を果たしている可能性が考えられる。 スーパーファミリーに属しており、同じTGF-スーパーファミリーに属するインヒビンのAサブユニットで構成される二量体構造をもっている。アクチビンは性腺以外にも、下垂体、神経系、骨髄、副腎、肝臓、膵などの様々な臓器での発現が知られており、局所において産生されオートクリン、パラクリン因子として細胞の分化や増殖および発生に大きな影響を及ぼしている。たとえばその作用として、赤芽球及び巨核球の分化誘導作用、神経栄養因子としての作用、またリンパ球、肝細胞、血管平滑筋細胞などの増殖制御因子としての作用が知られている。また発生段階では中胚葉の分化誘導因子としても働いている。このように、アクチビンAは発生の段階から分化・増殖をはじめとして様々な細胞調節機構に局所因子として関与する多機能性タンパク質である。そしてアクチビンAは、ラット膵においてラ氏島に特異的に発現しており、また胎生期の膵内分泌腺前駆細胞にも発現していることから、膵の分化および増殖において何らかの役割を果たしている可能性が考えられる。 AR42J細胞は化学物質アザセリンにより誘導されたラット膵腫瘍由来の培養細胞であり、正常膵外分泌腺、内分泌腺と同様に膵導管系上皮細胞に由来すると考えられている。またこの細胞はアミラーゼ、キモトリプシンなどの消化酵素を合成、分泌するという膵外分泌腺の特徴をもち、デキサメサゾンで処理することによりこれらの消化酵素の含有量が増加し膵外分泌腺としての特徴をより強く示すようになる。一方、このAR42J細胞の増殖はインスリン、ガストリン、EGFなどで促進されソマトスタチンで抑制される。これらのことより長年この細胞は膵外分泌腺細胞の増殖、分化および分泌機構の研究に多用されてきた。 AR42J細胞は、前述のようにデキサメサゾンによりさらに強く膵外分泌腺の方向に分化する。そしてアクチビンAをデキサメサゾンと同時に加えるとそのデキサメサゾンの作用は抑えられることを筆者らの研究グループは示してきた。つまり、アクチビンAはデキサメサゾンによるAR42J細胞のアミラーゼ含有量の増加を阻害するのである。さらにAR42J細胞をアクチビンA単独で処理すると、増殖は完全に停止しアミラーゼも消失する。このことは、アクチビンAがAR42J細胞を膵外分泌腺とは異なる方向へ分化させている可能性を示していると考えられる。また興味深いことに元来AR42J細胞はシナプス小胞のマーカーであるシナプトファイジンを発現し、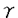 -アミノブチル酸などの神経伝達物質を放出するなど神経内分泌細胞の特徴も有する。AR42J細胞が膵発生の源である膵導管系上皮細胞由来の細胞であり、またアクチビンAが内分泌細胞や神経細胞などの様々な細胞の分化を修飾することを考えあわせれば、アクチビンAがAR42J細胞の神経内分泌細胞の特性に対し如何なる影響を与えるかは興味深い点であり、膵の発生および分化におけるアクチビンAの役割を考える上で重要であると考えられる。そこで本研究はAR42J細胞の形態と機能に対するアクチビンAの作用を明らかにする目的で行った。 -アミノブチル酸などの神経伝達物質を放出するなど神経内分泌細胞の特徴も有する。AR42J細胞が膵発生の源である膵導管系上皮細胞由来の細胞であり、またアクチビンAが内分泌細胞や神経細胞などの様々な細胞の分化を修飾することを考えあわせれば、アクチビンAがAR42J細胞の神経内分泌細胞の特性に対し如何なる影響を与えるかは興味深い点であり、膵の発生および分化におけるアクチビンAの役割を考える上で重要であると考えられる。そこで本研究はAR42J細胞の形態と機能に対するアクチビンAの作用を明らかにする目的で行った。 まず初めに免疫細胞化学的手法を用いて、アクチビンAによるAR42J細胞の形態的変化を検討した。AR42J細胞は4nMアクチビンA存在下で培養すると複数の突起を伸長し、一方アミラーゼ含有分泌顆粒は消失した。この伸長した突起は、神経に特異的な蛋白質であるであるニューロフィラメント160kDサブユニットと神経樹状突起に特異的な蛋白質MAP2を発現しており、このアクチビンAにより伸張したAR42J細胞の突起は神経突起に類似するものであると考えられた。またさらにアクチビンA処理をしたAR42J細胞では、シナプトファイジンの発現が増加していることがウェスタンブロッティングおよび免疫染色法で明らかになった。以上の結果からアクチビンAはAR42J細胞を神経内分泌細胞へ分化させると考えられた。 次にアクチビンAにより形態的に神経内分泌様細胞に分化したAR42J細胞の機能的変化を検討した。膵内分泌細胞は膵外分泌腺と同じく内胚葉由来の細胞であるにもかかわらず神経特異的エノラーゼやシナプトファイジンなどの神経特異的マーカーを発現し、特に膵ラ氏島B細胞はin vitroで培養するとニューロフィラメントを有する突起を伸長するなどの神経内分泌細胞の特徴を有する。これらの事実からアクチビンAによって神経内分泌細胞に分化したAR42J細胞が、膵ラ氏島B細胞の特徴を有する可能性が考えられたので、その機能的解析を電気生理学的に行った。まずAR42J細胞の電位依存性カルシウムチャネル(VDCC)の発現を調べるために、高カリウム細胞外液で細胞膜を脱分極させることによりVDCCを活性化させてその細胞内遊離カルシウム([Ca2+]c)に対する効果を検討したところ、[Ca2+]cの上昇はアクチビンA処理細胞において著しく増強していた。そこでVDCC電流量を測定したところ、アクチビンA処理および未処理のAR42J細胞の間にVDCC電流量の差を検出することはできなかった。しかしカフェインに対する[Ca2+]cの上昇反応がアクチビンA処理細胞にて著しく増強していた。カフェインは、カルシウム惹起性カルシウム放出機構(CICR)に関与する細胞内カルシウムプールから、カルシウムを放出をさせる働きがある。したがってこの結果はCICR機構がアクチビンA処理AR42J細胞において発達していることを示すものと考えられた。これらを考え合わせるとアクチビンA処理AR42J細胞での高カリウム細胞外液による細胞膜脱分極で惹起される[Ca2+]cの著明な上昇は、膵内分泌細胞で重要な働きを担っていると考えられているCICR機構がアクチビンA処理AR42J細胞で発達していることがその一つの要因であると考えられた。また、AR42J細胞に発現するVDCCの遺伝子解析でも、アクチビンA処理AR42J細胞の膵内分泌細胞様特徴が明らかになった。ラット膵ラ氏島B細胞からクローニングされ、インスリン分泌の調節を担っていると考えられている神経内分泌/B細胞特異的VDCC(B-VDCC)1サブユニットがアクチビンA処理AR42J細胞に発現していることがRT-PCRで明らかになった。このB-VDCC1サブユニットはインスリン分泌培養細胞すべてに発現している訳ではなく、RINm5FやTC-3には発現しているがHIT-T15には発現していない。このことは、B-VDCC1サブユニットを発現していることは、アクチビンA処理AR42J細胞が膵ラ氏島B細胞様の特徴を潜在的に備え持っていることを示唆すると考えられた。 アクチビンA処理AR42J細胞の膵ラ氏島細胞様の特徴をさらに明らかにするため、そのトルブタマイドに対する反応を検討した。トルブタマイドは経口糖尿病治療薬として広く使用されているスルフォニル尿素製剤の一つであり、膵ラ氏島B細胞においてKATPチャネルまたはスルフォニル尿素受容体と結合し、KATPチャネルの活性を抑制することで細胞膜を脱分極させ、その結果VDCCを開いて[Ca2+]cを上昇させる。アクチビンA処理AR42J細胞ではトルブタマイドに反応して[Ca2+]cが上昇し、かつパッチクランプ法でもKATPチャネルの存在が確認された。これらから、アクチビンA処理AR42J細胞には、膵ラ氏島内分泌細胞に特徴的な二種類のチャネルB-VDCCとKATPチャネルが発現しており膵内分泌細胞類似の電気生理学的特徴を有すると考えられた。 以上のように本研究において、アクチビンAはAR42J細胞を形態的に神経内分泌細胞に分化させ、その分化した細胞は膵内分泌細胞類似の機能的特徴を有することが明らかになり、アクチビンAが膵の分化誘導因子である可能性が示唆された。またAR42J細胞がデキサメサゾンで膵外分泌細胞に分化し、一方アクチビンAで神経内分泌細胞に分化することから鑑みて、AR42J細胞は膵の外分泌腺および内分泌腺への分化を研究するうえでのモデル系になり得ると考えられた。 | 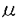 M)刺激により細胞内カルシウム濃度が上昇し、未処理細胞では上昇しなかった。また、アクチビンA処理したAR42J細胞ではATP感受性カリウムチャネル(K-ATPチャネル)が発現していることがパッチクランプ法で示された。
M)刺激により細胞内カルシウム濃度が上昇し、未処理細胞では上昇しなかった。また、アクチビンA処理したAR42J細胞ではATP感受性カリウムチャネル(K-ATPチャネル)が発現していることがパッチクランプ法で示された。