本研究はB細胞や好酸球の分化増殖に関与しているインターロイキン-5の発現分子機構の解析を試みたのもであり、下記の結果を得ている。 1.IL-5発現細胞株であるATL16Tを用いて、IL-5プロモーター上に転写活性能を有する20塩基からなるエレメント(G1領域)を絞り込んだ。ATL-16Tの核抽出液を用いてゲルシフト解析を行い、そのエレメントに結合するタンパク質の存在を明らかにした。このタンパク質を単離する目的で、ATL-16Tの発現ライブラリーを、サウスウエスタン法を用いてスクリーニングし、GATAファミリータンパク質がこのエレメントに結合する事を明らかにした。 2.各GATAファミリータンパク質(GATA-2、GATA-3、及びGATA-4)に対する抗体を作製した。ATL-16T細胞核抽出液中に存在するGATAファミリータンパク質の内、GATA-4が主にG1領域に結合することを明らかにした。また、組み替え発現させた各種GATAファミリータンパク質を用いてG1領域に対する結合親和性をGATA-2、GATA-3、及びGATA-4に関して比較したところ、GATA-4が最もG1領域に結合親和性が強いことを明らかにした。 3.G1領域に結合するタンパク質がGATA-4であることが明らかとなった時点で、Jurkat細胞を用いた形質導入実験を行った。IL-5プロモーターの活性化にはGATA-4とTPA-A23187刺激の両方が必要であり、いずれか一方ではIL-5プロモーターは活性化されないことを示した。また、IL-5プロモーターに変異を入れた実験より、-70に存在するGATAエレメントと、-45に存在するCLEOエレメントの2つが、IL-5プロモーターの活性化に必要なエレメントであることを明らかにした。 4.ヒトGATA-4のcDNAをクローニングし、その全塩基配列を決定した。 以上,本論文はGATA-4のクローニング、及び形質導入実験によりIL-5プロモーター領域の解析を行い、IL-5の発現分子機構を明らかにしたものである。本研究は,これまでほとんど解析の進んでいなかったIL-5の、アレルギー疾患における関与の解明に重要な貢献をなすと考えられ,学位の授与に値すると考えられる. | 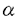 -GATA4)をCOS細胞に形質導入して発現させ、その抽出液を用いてゲルシフト解析を行った。各変異を入れたラベルしていないオリゴヌクレオチドによる競合実験において、COS細胞で発現させた組み換え型GATA-4は、ATL-16T細胞核抽出液に存在する天然型GATA-4と同様の結合様式を示した。すなわちAGATAG配列に変異を入れたオリゴヌクレオチドでは結合は阻害されず、それ以外の部分に変異を入れたオリゴヌクレオチドでは阻害された。このことから、GATA-4は実際にG1領域のAGATAG配列に結合することが確認された。次に、GATA-4がIL-5プロモーターを実際に活性化するかについて検討を行った。IL-5非産生T細胞株であるJurkat細胞に、IL-5プロモーター領域をルシフェラーゼ遺伝子の上流につないだプラスミド(pUCIL5-Luci)と、pSSR
-GATA4)をCOS細胞に形質導入して発現させ、その抽出液を用いてゲルシフト解析を行った。各変異を入れたラベルしていないオリゴヌクレオチドによる競合実験において、COS細胞で発現させた組み換え型GATA-4は、ATL-16T細胞核抽出液に存在する天然型GATA-4と同様の結合様式を示した。すなわちAGATAG配列に変異を入れたオリゴヌクレオチドでは結合は阻害されず、それ以外の部分に変異を入れたオリゴヌクレオチドでは阻害された。このことから、GATA-4は実際にG1領域のAGATAG配列に結合することが確認された。次に、GATA-4がIL-5プロモーターを実際に活性化するかについて検討を行った。IL-5非産生T細胞株であるJurkat細胞に、IL-5プロモーター領域をルシフェラーゼ遺伝子の上流につないだプラスミド(pUCIL5-Luci)と、pSSR