細胞が伸展されると、その機能が影響を受けることが多くの細胞で知られている。培養骨格筋細胞において、伸展刺激により活性化されるチャネル(機械受容チャネル)の存在が、1984年にGuharay&Sachsにより、電気生理学的手法によって初めて示唆された。現在では、原核生物から哺乳類まで、卵細胞、赤血球、内耳有毛細胞、神経細胞、心筋細胞、平滑筋細胞、腎上皮細胞、血管内皮細胞など広く様々な細胞において、この機械受容チャネルの存在が示されており、伸展刺激が細胞機能に影響を与える機構の重要な鍵を握る分子であると考えられている。 伸展刺激により平滑筋が収縮することは、1947年にBozlerらにより示されているが、その詳しいメカニズムについては不明であり、また定量的な解析も十分には行われていなかった。子宮は、妊娠経過とともに伸展が加わり、ダイナミックに変化する特徴を持つ臓器である。その収縮制御機構を考える上で、伸展刺激の及ぼす影響を明らかにすることは、分娩発来機構の解明など、生理学的にも臨床的にも重要な意味を持つと思われる。そこで、細胞内Ca2+濃度と発生張力を同時測定するとともに、筋標本の長さを自由にコントロールして伸展刺激を与えることのできる方法を開発し、伸展刺激の子宮筋収縮制御機構に対する影響を明かにする目的で研究を行った。 標本は、妊娠時と非妊娠時のラットの子宮から縦走筋を、長さ約1mm,幅200-300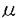 mの筋束標本として、実体顕微鏡下で作成した。あらかじめ、Ca2+蛍光指示薬であるFura-2 AMを標本に負荷し、標本の一端を張力トランジューサーに取付けて等尺性張力を測定した。対側は、コンピューター制御下のサーボモーターに取付け、あらかじめ設定した速さで、標本の長さを変化させ、定量的な伸展刺激を与えた(図1)。同時に細胞内に導入したFura-2の蛍光強度変化から細胞内Ca2+濃度を測定した。また、筋標本の初期長は、生体内の長さに対応するように、あらかじめ静止張力を与え、非妊娠時では、弛緩長の125%を初期長とした。また、測定はすべて30℃で行った。 mの筋束標本として、実体顕微鏡下で作成した。あらかじめ、Ca2+蛍光指示薬であるFura-2 AMを標本に負荷し、標本の一端を張力トランジューサーに取付けて等尺性張力を測定した。対側は、コンピューター制御下のサーボモーターに取付け、あらかじめ設定した速さで、標本の長さを変化させ、定量的な伸展刺激を与えた(図1)。同時に細胞内に導入したFura-2の蛍光強度変化から細胞内Ca2+濃度を測定した。また、筋標本の初期長は、生体内の長さに対応するように、あらかじめ静止張力を与え、非妊娠時では、弛緩長の125%を初期長とした。また、測定はすべて30℃で行った。 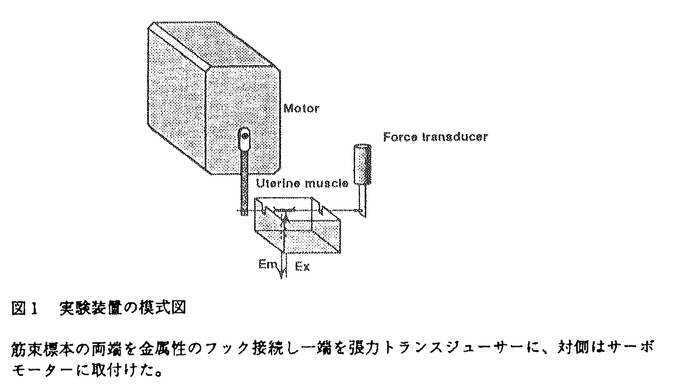 図1 実験装置の模式図筋束標本の両端を金属性のフック接続し一端を張力トランスジューサーに、対側はサーボモーターに取付けた。 図1 実験装置の模式図筋束標本の両端を金属性のフック接続し一端を張力トランスジューサーに、対側はサーボモーターに取付けた。 筋束標本にある大きさ以上の伸展刺激を加えると、まず細胞内Ca2+濃度の上昇が起こり、それに続いて収縮が認められた。この反応は、伸展刺激の大きさと速度に依存して起こり、ある閾値を持って反応した。0.5秒間で伸展刺激を与えた場合、(図2A)に示した標本では、0.5秒間で初期長の15%の伸展刺激を与えても、細胞内Ca2+の上昇も収縮も起こらなかった。しかし、20%の伸展刺激では細胞内Ca2+の上昇と収縮が起こり、Ca2+の上昇値は、25%の伸展刺激でも、20%の場合と同じであった。また、(図2B)に示すように、20%の伸展刺激でも、0.5秒間で与えた場合は細胞内Ca2+の上昇と収縮が起こったが、10秒間かけゆっくり伸展した場合には受動的な張力の発生以外には反応はみられなかった。このように伸展刺激は、伸展刺激の大きさと速度の変化に対して、「全か無か」の細胞内Ca2濃度変化を示した。この結果から、伸展刺激による反応にはCa2+スパイク(活動電位)の関与が示唆された。 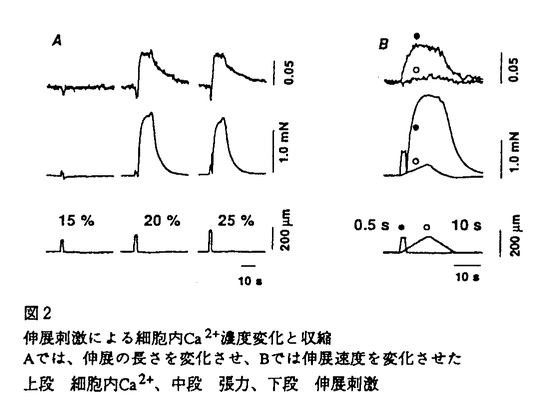 図2 伸展刺激による細胞内Ca2+濃度変化と収縮Aでは、伸展の長さを変化させ、Bでは伸展速度を変化させた上段 細胞内Ca2+、中段 張力、下段 伸展刺激 図2 伸展刺激による細胞内Ca2+濃度変化と収縮Aでは、伸展の長さを変化させ、Bでは伸展速度を変化させた上段 細胞内Ca2+、中段 張力、下段 伸展刺激 次に、伸展刺激がどのような機序を介して、平滑筋の収縮を起こすかを調べる目的で以下の実験を行った。細胞外のCa2+を除去してEGTAを投与したり、Ca2+拮抗薬であるニカルジピン10nMを投与すると、伸展刺激に対する反応は完全に抑制された。しかし、細胞内Ca2+ストアのCa2+-ATPase阻害薬であるサイクロピアゾン酸10Mにより、ストアからのCa2+放出を完全に抑えてもこの反応は抑制されなかった。また、アトロピン1Mやテトロドトキシン1Mでも抑制されなかった。従ってこの反応は、細胞内Ca2+ストアからのCa2+放出や自律神経の活動には依存せず、筋細胞が直接伸展刺激に応答し、細胞外からのCa2+流入により起こる筋原性の収縮であることが示された。 さらに、オキシトシンによる律動的収縮に対する、伸展刺激の影響について調べた。子宮平滑筋は、オキシトシンなどのアゴニストにより、律動的な細胞内Ca2+上昇とそれに伴う収縮を起こす。伸展刺激を、収縮と収縮との間の弛緩期に与えると、本来の収縮のリズムより早く、次の細胞内Ca2+濃度の上昇と収縮が起こったが、リズムそのものには大きな影響はなかった。即ち律動的な収縮のリズムの位相が早められた。また、個々の律動的収縮の収縮時間が延長し、伸展刺激はオキシトシンによる収縮を増強することが明かとなった。 以上の結果は、機械受容チャネルが子宮筋においても存在し、伸展刺激はこのチャネルを活性化して細胞膜の脱分極を起こし、それがある閾値に達すると、Ca2+スパイクが生じて収縮に至ることを示唆している。さらに、伸展刺激はオキシトシンによる律動的収縮を強めることもわかった。このように、機械刺激は子宮筋の収縮制御に重要な役割を果たしている。機械受容チャネルに関する研究は、これまで単離細胞や培養細胞を用いて主に行われてきた。本研究では、筋束標本を用いたことにより、生体に近い条件において、伸展刺激が子宮平滑筋の収縮制御に関わっていることを、細胞内Ca2+と張力とを同時測定することで、明かにした。子宮は妊娠の経過とともに胎児による伸展刺激を受ける。本研究によって示された結果から、このような伸展は子宮筋がオキシトシン等の刺激に対して強く反応する準備状況をつくり出すのに役立つことがわかってきた。 |