血管新生は、正常個体においては胎生期における循環器系の形成や多くの臓器の構築に重要な役割を果たし、成人では女性の性周期における黄体形成、子宮内膜の一過性の増殖にも関与する。病的状態としては、炎症や創傷治癒過程、糖尿病網膜症による眼底病変や関節リウマチなどにおいても血管新生は深く関与している。また、腫瘍血管新生が固形腫瘍の増殖に必須であることも明らかにされている。このように、血管新生は多くの生物学的過程において重要な役割を果たしている。近年、血管新生に密接に関与する増殖因子とその受容体が明らかになってきた。その代表として、VEGF(Vascular Endothelial Growth Factor)と、その受容体であるFlt-1(Fms-like tyrosine kinase)、Flk-1(Fetal liver kinase)が注目を浴びている。このVEGF/Flt-1,Flk-1 systemが内皮細胞制御の中心的機構を担っていることが示唆され始めている。 VEGFは、分子量約45KDaのヘパリン結合性の蛋白で、二量体を形成している。mRNAのスプライシングの違いにより4つのサブタイプ(VEGF121,VEGF165,VEGF189,VEGF206)が存在し、いずれの型もシグナルペプチドをもち、細胞外に分泌される。しかし、VEGF189とVEGF206にのみ存在するexon 6は塩基性アミノ酸に富んでおり、細胞膜表面のヘパリン様物質に結合する性質がある。 VEGFと類似性を示す因子としてPIGF(Placenta Growth Factor)が近年単離され、VEGFと高い相同性を示している。mRNAのスプライシングの違いにより、PIGF131(PIGF-1)、PIGF152(PIGF-2)の2つのサブタイプが現在までに報告されており、VEGF同様のexon 6を有しているのはPIGF-2のみである。 一方、VEGFの受容体と考えられているFlt-1は渋谷正史らによりヒト胎盤cDNAライブラリーより単離された受容体型チロシンキナーゼで、fms遺伝子群と構造類似性を示すことよりfms-like tyrosine kinase(Flt-1)と名付けられた。Flk-1もFltファミリーに属し、これら遺伝子の発現は内皮細胞系にほぼ限局している。増殖因子の生体内での結合部位を同定することは、その因子の作用部位を同定し、生物学的意義を理解する上で大変重要である。内皮細胞増殖促進作用を有するVEGFと、内皮細胞特異的に発現しているFlt-1,Flk-1との真の結合部位、また、それら受容体の局在部位の相違を可視化する簡便な方法として、筆者はラットおよびマウス胎仔組織と放射性ヨード標識されたVEGF蛋白(以後、125I-VEGF)を用いての結合実験を行った。実験方法として、正常ラット(1週令〜9週令)より各々臓器を摘出、凍結し、クライオスタットにて約6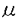 m-8mの厚さで凍結組織切片を作成した。マウス胎仔からも同様に凍結組織切片を作成した。この凍結切片をDME(Dulbecco Modified Eagle Medium;10% FCS,25mM HEPES,0.5mM MgCl2,4g/ml leupeptin,5ng PMSF添加)にて3時間反応し、その後、同溶液に12.5pM〜150pMの125I-VEGFを加え、さらに3時間反応した。非特異的結合を除外するため、各々の連続切片にて125I-VEGF溶液中に0.2nM〜4nMの未標識VEGFを加えて結合抑制をみた。また、他の増殖因子であるPDGF、bFGF、EGFを加えての競合阻害実験も行った。0.2nM〜4nM PIGFによる結合阻害によりFlt-1とFlk-1の存在比についても検討した。また、結合実験で用いた凍結組織切片で内皮細胞を同定するため、内皮細胞マーカーであるvon Willebrand Factorを免疫組織化学的に染色した。 m-8mの厚さで凍結組織切片を作成した。マウス胎仔からも同様に凍結組織切片を作成した。この凍結切片をDME(Dulbecco Modified Eagle Medium;10% FCS,25mM HEPES,0.5mM MgCl2,4g/ml leupeptin,5ng PMSF添加)にて3時間反応し、その後、同溶液に12.5pM〜150pMの125I-VEGFを加え、さらに3時間反応した。非特異的結合を除外するため、各々の連続切片にて125I-VEGF溶液中に0.2nM〜4nMの未標識VEGFを加えて結合抑制をみた。また、他の増殖因子であるPDGF、bFGF、EGFを加えての競合阻害実験も行った。0.2nM〜4nM PIGFによる結合阻害によりFlt-1とFlk-1の存在比についても検討した。また、結合実験で用いた凍結組織切片で内皮細胞を同定するため、内皮細胞マーカーであるvon Willebrand Factorを免疫組織化学的に染色した。 腎臓の切片では125I-VEGFの結合は皮質に存在する糸球体で最も強く、髄放線や乳頭周辺にも結合像がみられた。また、同部位の血管内皮細胞にvon WillebrandFactorの免疫染色も強く陽性であった。眼球の切片では、網膜周辺部、虹彩、毛様体の血管に沿って結合像が認められ、その内皮細胞においてvon Willebrand Factorの免疫染色は陽性だったが、正常では血管系の存在しない角膜や水晶体、硝子体には結合像は見られなかった。肺の切片では肺胞の虚脱した状態にて125I-VEGFの結合は肺胞中隔の毛細血管及び他の血管壁に認められ、von Willebrand Factorの免疫染色は、肺胞中隔毛細血管以外の血管壁に沿って陽性であった。脳の切片ではmicrovesselに125I-VEGF結合像がみられ、同部位にvon Willebrand Factorの免疫染色も陽性であった。肝臓の切片では、125I-VEGFは小葉間動静脈や中心静脈、類洞等の血管壁に結合するため肝臓の切片は全体的に、び漫性に結合像を呈したが、小葉間動静脈や中心静脈の血管壁により強く結合していた。von Willebrand Factorの免疫染色は、類洞壁内皮細胞以外の血管壁の内皮細胞に陽性像を呈した。また、マウス胎仔において、125I-VEGF結合は脳軟膜、網膜周辺部、背部脂肪組織、肺、肝臓、腎臓等に認められた。 以上のように、今回の実験結果によると、125I-VEGFの結合部位は、ラット正常組織およびマウス胎仔において広く分布し、この分布パターンは血管系の分布パターンとほぼ一致している。そして、このVEGFの結合部位はすなわち、VEGFの主な受容体であるFlt-1、Flk-1の局在部位と考えられる。VEGFはヘパリン親和性蛋白であることより、細胞膜上のヘパリン様物質や基底膜等に結合している可能性を否定するために、競合阻害実験としてPDGF,bFGF,EGFの増殖因子を加えたが、これらの因子では結合は阻害されなかった。また、VEGFそのものによる競合阻害実験では結合は抑制されたことより、この125I-VEGFの結合は非特異的結合ではなく、その受容体との結合そのものを示していると示唆された。 VEGF受容体の報告によると、VEGFはFlt-1、Flk-1両者に結合するのに対し、PIGFはFlt-1にのみ結合可能である。この二つの蛋白の結合パターンの違いを用いて受容体の分布の相違を見るため、125I-VEGF結合に対するPIGFによる競合阻害実験を行った。PIGFは125I-VEGF結合を部分的に阻害し、この阻害効果はすなわち、PIGFとFlt-1の結合によるものと考えられた。これらを定量化した結果、VEGF受容体はFlt-1:Flk-1=約1:1の割合で存在することが示唆された。 125I-VEGFによる結合実験はリガンドとレセプターとの結合部位を組織切片上で確認し、作用部位を同定できる優れた方法である。リガンドに対する受容体が同定されている場合、受容体のmRNAの発現を検出するin situ hybridization法や、蛋白レベルでの発現を検出する免疫組織化学があるが、いずれも受容体が機能分子として細胞膜上に発現されていることを確認することはできない。今回行った、放射性ヨード標識された蛋白を用いたLigand-Receptor Binding AssayはVEGF受容体の局在部位やその量、存在比などを解析し、生体内で実際に作用している部位を同定するため有用であると考えられた。 |