形態形成は、細胞の空間的な再配列や、細胞同士の選択的な接着や剥離、協調性を保った細胞の分化と増殖により生ずる高度な生命現象である。このような形態形成に関わる分子メカニズムを明らかにするため、私は、活性分子であり、さらに蛋白を切断することにより大きく構造や活性を変化させるproteaseに注目し、形態形成におけるproteaseの機能を明らかにすることを試みた。 成虫原基は、完全変態昆虫の幼虫内に存在する未分化な成虫組織であり、蛹の時期に変態ホルモンecdysoneの刺激により成虫組織へと分化する。当教室では、成虫原基を幼虫体内から摘出し、ecdysone存在下で培養することにより成虫組織への発生がin vitroにおいて再現されている。修士課程において、成虫原基のin vitro発生系を用い蛋白内のprolineのC末端側を水解する酵素であるprolyl endopeptidaseが、成虫原基の発生初期の形態形成に関与していることを見出し、この蛋白をセンチニクバエ胚由来培養細胞より精製した。 本研究において私は、センチニクバエprolyl endopeptidaseのcDNA cloningを行い構造の解析を行った。また、胚及び成虫原基の発生におけるprolyl endopeptidaseの機能を解析した結果、prolyl endopeptidaseが形態形成の初期に必須な細胞分裂に重要な役割を果たしていることを明らかにした。 1.センチニクバエprolyl endopeptidaseのcDNA cloning センチニクバエprolyl endopeptidaseの一次構造を明らかにし、その遺伝子発現を解析するために、センチニクバエprolyl endopeptidaseのcDNAを、精製の出発材料であるセンチニクバエ胚由来培養細胞NIH-Sape-4のcDNA libraryより単離した。得られたcDNAは2514 bpのインサートを含み、714個のアミノ酸からなるオープンリーディングフレームを有していた。予想されるアミノ酸配列中には、決定したprolyl endopeptidaseの部分アミノ酸配列がすべて含まれ、さらに得られたcDNAを大腸菌に発現させて調製したrecombinantはnative prolyl endopeptidaseと同等の活性を有していた。これらの結果から、このクローンはセンチニクバエprolyl endopeptidaseのcDNAであることが示された。他の蛋白との相同性を検索した結果、哺乳類と細菌類のすでにcDNAが単離されている3種のprolyl endopeptidaseと高い相同性を有していた。 2.センチニクバエprolyl endopeptidase mRNAの胚及び成虫原基発生の際の発現 胚及び成虫原基の発生の際のセンチニクバエprolyl endopeptidaseのmRNAの発現をNorthem blot解析により検出した。その結果、センチニクバエprolyl endopeptidaseのmRNAは未受精卵中に検出され、matemal messageとして貯蔵されていることが明らかとなった。また、胚発生時においては、発生初期にmRNA量が最大となった。これらのことは、センチニクバエprolyl endopeptidaseが胚発生において受精直後からの極めて初期の過程に重要な役割を果たしていることを示唆している。また、成虫原基の発生においても、未発生から発生初期(eversion)の原基でmRNA量が最大であり、その後、減少した。これは、成虫原基の発生においてもセンチニクバエprolyl endopeptidaseが発生初期に働いていることを示唆している。また、この結果は、修士課程において示した、prolyl endopeptidase阻害剤が成虫原基の発生初期のみを特異的に阻害する結果とよく一致する。 3.胚由来培養細胞NIH-Sape-4におけるprolyl endopeptidaseの機能の解析 胚発生において受精直後の初期胚の内部では、急激な細胞分裂が行われ、さらに、この時期にprolyl endopeptidaseの発現が最大になることから、prolyl endopeptidaseは胚の細胞増殖に関与する可能性が考えられる。そこで、胚発生におけるprolyl endopeptidaseの機能を解析するために胚由来培養細胞NIH-Sape-4の増殖に対するprolyl endopeptidase特異的阻害剤(ZTTA:carbobenzoxy-thioprolyl-thioprolyl-acetal)の効果を調べた。その結果、NIH-Sape-4の培養系にZTTAを添加すると、容量依存的に培養細胞の増殖が阻害された。さらにこのときのDNA複製を3H-thymidineのDNAへの取り込みにより解析した結果、ZTTAにより細胞増殖と同様に取り込みが阻害された。一方、蛋白合成やecdysoneによる細胞形態の変化はZTTA添加によっても全く影響を受けなかった。これらの結果は、prolyl endopeptidaseが培養細胞の増殖、特にDNA複製に関わっていることを示しており、また、prolyl endopeptidaseは胚発生の初期の形態形成に重要な細胞分裂に関わっている可能性が考えられる。 4.培養細胞及び初期胚内のprolyl endopeptidaseの局在 prolyl endopeptidaseの胚発生における機能をさらに検討するために、胚由来培養細胞NIH-Sape-4内のprolyl endopeptidaseの局在を明らかにした。抗体は、immunoblotにおいて培養細胞lysate中prolyl endopeptidaseのみを特異的に検出するものを用いた。その結果、prolyl endopeptidaseは、核の内部に局在していた。さらに、共焦点レーザー顕微鏡を用い細胞内の局在を詳細に検討した結果、prolyl endopeptidaseは、核小体と考えられる核内のDNAが存在しない部位の上下に認められた。また、Iog phase及びfull growthの細胞内のprolyl endopeptidaseを検出した結果、full growthの細胞はlog phaseのものと比較して、核内の蛍光が減少、消失していた。これらの結果は、prolyl endopeptidaseは、培養細胞の核内において細胞の増殖に関わるなんらかの機能を担っていることを示している。また、初期胚においても同様にprolyl endopeptidaseの局在を検討した結果、培養細胞と同様に核内に局在していることが明らかとなった。この結果から、prolyl endopeptidaseは、初期胚においても培養細胞と同様の機能を有していることが明らかとなった。 5.成虫原基の形態形成におけるprolyl endopeptidaseの機能の解析 成虫原基の発生において、prolyl endopeptidaseの遺伝子発現が胚発生と同様の挙動を示したことから、prolyl endopeptidaseは、成虫原基の発生においても細胞増殖に関与している可能性が考えられた。そこで、成虫原基の発生に従って、細胞増殖がどのように変化するかを検討した結果、これまで、主に細胞の移動と細胞容積の増大により行われると考えられたが、発生初期にのみ細胞増殖が存在することを見出した。成虫原基の発生のprolyl endopeptidase阻害剤による阻害は、この細胞増殖が存在する初期のみ認められたことから、胚発生と同様に、prolyl endopeptidaseは発生初期の細胞増殖に働いている可能性が考られる。実際、この発生初期の成虫原基での3H-thymidineの取り込みはZTTAにより、容量依存的に阻害された。このとき、ZTTAが成虫原基の細胞増殖を阻害しはじめる濃度(100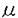 M)は成虫原基の発生を阻害する濃度と一致した。このことは、成虫原基においても胚発生と同様に形態形成に先立つ細胞増殖が必要であり、prolyl endopeptidaseは、この細胞増殖に働いていることを示している。 M)は成虫原基の発生を阻害する濃度と一致した。このことは、成虫原基においても胚発生と同様に形態形成に先立つ細胞増殖が必要であり、prolyl endopeptidaseは、この細胞増殖に働いていることを示している。 6.まとめと考察 私は、胚及び成虫原基の発生に関与しているprolyl endopeptidaseのcDNAをcloningし、その一次構造を明らかにした。さらに、胚及び成虫原基の発生におけるprolyl endopeptidaseのmRNAの発現を見出した結果、prolyl endopeptidaseはmatemal messageとして未受精卵に存在し、胚発生と成虫原基発生の初期に発現量が最大になることを見出した。また、prolyl endopeptidaseは胚由来培養細胞の増殖と、成虫原基の発生初期に行われる細胞増殖に関わっており、特に培養細胞と初期胚においては核内に局在していることを示した。 胚発生と成虫原基の発生においては、まず細胞増殖により細胞数が増大し、その後、細胞の移動と細胞容積の増大がおきる。prolyl endopeptidaseは形態形成に不可欠な初期の細胞増殖に働いている。さらに、昆虫の胚発生においては受精直後は細胞分裂を伴わない核分裂のみが起きることから、prolyl endopeptidaseは細胞周期のみならず、DNA複製に直接働いている可能性も考えられる。本研究は、核内のproteaseが組織レベルの形態形成とそれに伴う細胞分裂に働いていることを示すものであり、proteaseとdevelopmentの研究分野に新たな視点を与えるものである。 |