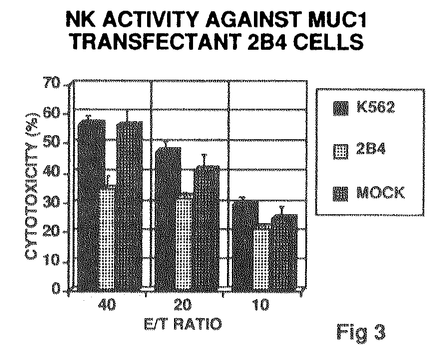
本論文は、細胞表面ムチンのグリコシレーション(糖蛋白の糖鎖部分の生合成)の多様性を明らかにし、さらにグリコシレーションがどのように制御されているかを、分子細胞生物学的な手法を駆使して解明した結果をまとめたものである。細胞表面にレクチン(糖鎖結合分子)によって認識される糖鎖が表出している場合しばしばムチンという形をとる。細胞表面のムチンと内在性レクチンとの相互作用は、癌細胞による転移形成、白血球の炎症部位への集積などの生体内の細胞交通を制御するメカニズムの分子レベルでの基盤となっている。ムチンとは多くのセリン/スレオニン結合型糖鎖をもつ高分子量の糖蛋白であり、その分布は臓器によりまた細胞の種類により極めて特徴的である。ムチンのコア蛋白の発現及びムチンに含まれる糖鎖構造の発現は極めて多様で、細胞種に特異的であると考えられいる。ムチンの生合成において蛋白部分と糖鎖部分とが独立に制御されているのかあるいは糖鎖の発現がコア蛋白質部分の構造によって左右されているのかは明らかになっていなかった。 学位申請者は上皮性ムチンのひとつであるMUC1ムチンに提示される糖鎖抗原に着目し、ヒト培養細胞のムチンあるいは白血病細胞へのMUC1遺伝子導入により得たMUC1ムチン安定発現細胞上のMUC1ムチンについて糖鎖抗原の構造を糖鎖特異的モノクローナル抗体やレクチンの結合性に基づいて検討した。また、MUC1ムチンの機能を探る試みのひとつとして、自然免疫を担うナチュラルキラー(NK)細胞の細胞障害性に対するMUC1ムチン発現細胞の感受性について検討した結果も示されている。 第一部では、一種類のムチンつまりこの場合はMUC1遺伝子産物をコアに持つムチンが、複数種の細胞株で発現した場合、それらのグリコシレーションがどのように異なるかについて検討されている。糖鎖の検出には、モノクロナル抗体と植物レクチンとが用いられた。三種の上皮性細胞株の産生するMUC1ムチンはいずれもレクチン結合のパターンが異なっており、MUC1ムチンが提示する糖鎖構造は発現する細胞により異なることが判明し、それぞれの細胞のもつ糖転移酵素に依存すると考えられた。次にひとつの細胞が複数種のムチンを発現している場合について検討した。レクチン及びモノクロナル抗体を駆使した実験結果より、ひとつの細胞がMUC1とMUC2ムチンを発現した場合それらの間で主要な糖鎖構造が異なるという現象が明らかになった。 第二部では、ヒト赤白血病細胞株K562にMUC1遺伝子を導入しMUC1ムチン安定発現クローンを得て、内在性のムチン様分子の一つであるロイコシアリン(CD43)と強制発現されたMUC1ムチンとの間でグリコシレーションの違いがあるかどうか査定した。レクチンやモノクロナル抗体を用いた検出の結果、強制発現されたMUC1ムチンではシアル化の度合の低い糖鎖がCD43に比較すると主要部分を占めていることが確かめられた。MUC1ムチンは細胞内でのプロセシングの速度がCD43よりも速いことがその原因として示唆され、ムチンの糖鎖構造がコアペプチドによって強い影響を受けることが明らかになった。 第三部では、ヒト赤白血病細胞にMUC1ムチン遺伝子を導入した際に、この細胞のNK細胞に対する感受性にどのような変化が起こっているかを査定した。MUC1ムチン発現クローンの細胞障害性は親株細胞および対照株に比べ有意に低下していた。MUC1ムチン発現細胞をシアリダーゼ処理して糖鎖末端のシアル酸を除去した後、あるいはN-アセチルガラクトサミンのベンジルグリコシドで処理しO-結合型糖鎖の伸長を阻害した後NK細胞に対する感受性を測定したが、NK感受性のパターンに大きな変化はみられなかったことから、MUC1ムチン全体の構造がNK細胞との相互作用に影響を与えるためと推定された。 要約すると、MUC1ムチンと他のムチンとを同一の細胞が産生している際にそれぞれの糖鎖構造が異なることが確認された。また、MUC1ムチンがシアリルTn抗原、Tn抗原、T抗原など腫瘍関連抗原を提示しうることが確認された。MUC1ムチンの遺伝子導入による強制発現が神細胞に対する抵抗性を産み出すことが示され、担癌生体が癌細胞の排除をしようとするメカニズムを阻害している可能性が示唆された。 以上の様に、学位申請者名取修は、非特異的及び特異的な癌に対する免疫応答の際の認識分子として重要なMUC1ムチンのグリコシレーションの制御機構と、MUC1ムチンを発現した細胞の非特異的自然免疫に対する感受性に関する新しい知見を得た。これらの研究成果は腫瘍生物学及び免疫学に資するところが大であり、学位申請者は博士(薬学)の学位を受けるにふさわしいと判断した。 | 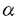 2-3Gal
2-3Gal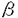 1-4GlcNAcに結合)が結合したがMUC2ムチンには結合しなかった。一方MUC2ムチンにはACA(NeuAc
1-4GlcNAcに結合)が結合したがMUC2ムチンには結合しなかった。一方MUC2ムチンにはACA(NeuAc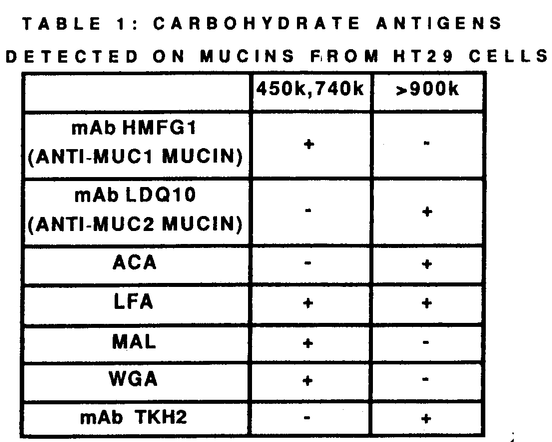
 g/mlのgeneticinによりセレクションを行い、限界希釈法によりクローン化を行った。クローン化した細胞表面にMUC1ムチンやムチン関連糖鎖抗原が発現しているかどうか確かめるため、当研究室で樹立したシアル酸を含む成熟型糖鎖をもつMUC1ムチンを認識するmAb MY.1E12あるいはMUC1ムチンコアペプチドを認識するmAbHMFG2を用いたフローサイトメトリー法(FCM)およびwestern blot法による解析を行った。[3H]グルコサミンを30分取り込ませた後一定時間培養した細胞の可溶化物をMY.1E12あるいは抗CD43 mAb DFT1で免疫沈降しSDS電気泳動後オートラジオグラフィーを行った。
g/mlのgeneticinによりセレクションを行い、限界希釈法によりクローン化を行った。クローン化した細胞表面にMUC1ムチンやムチン関連糖鎖抗原が発現しているかどうか確かめるため、当研究室で樹立したシアル酸を含む成熟型糖鎖をもつMUC1ムチンを認識するmAb MY.1E12あるいはMUC1ムチンコアペプチドを認識するmAbHMFG2を用いたフローサイトメトリー法(FCM)およびwestern blot法による解析を行った。[3H]グルコサミンを30分取り込ませた後一定時間培養した細胞の可溶化物をMY.1E12あるいは抗CD43 mAb DFT1で免疫沈降しSDS電気泳動後オートラジオグラフィーを行った。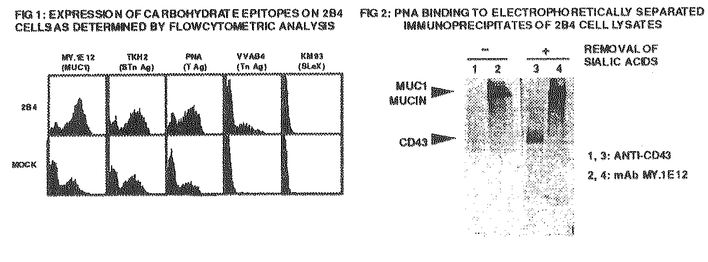
 -カウンターで測定した。細胞障害性は以下の式により算出した。細胞障害性(%)=(サンプル放射活性-自然放射活性)×100/(最大放射活性-自然放射活性)
-カウンターで測定した。細胞障害性は以下の式により算出した。細胞障害性(%)=(サンプル放射活性-自然放射活性)×100/(最大放射活性-自然放射活性)