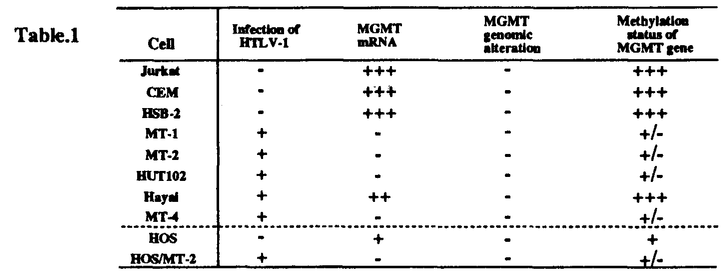
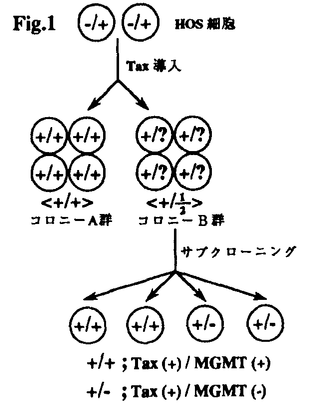
ヒトT細胞白血病ウイルス(HTLV-1)は、成人T細胞白血病(ATL)の原因ウイルスである。とりわけ、HTLV-1がコードするTax蛋白質は、白血病生成に大きく関わると考えられている。しかし、一方でTaxの直接作用だけではATL発症に充分ではないことが示唆され、HTLV-1感染後に細胞の遺伝子発現の変化が必要であることが想定されている。この論文は、ほとんど全てのHTLV-1感染細胞株で、DNA修復遺伝子であるO6-メチルグアニンーDNAメチルトランスフェラーゼ(MGMT)遺伝子の発現が検出されないという新しい知見を見い出し、MGMT遺伝子発現抑制機構の解析を行なった結果、HTLV-1がコードするTax蛋白質の今までに知られていなかった新しい作用の存在を明らかにした点で評価できる。 この論文では、まず、ATL細胞が高頻度の染色体異常を持つという知見に着目し、ゲノムの安定性への関与が知られているDNA修復遺伝子の発現異常をHTLV-1感染細胞で検討した。その結果、ほとんど全てのHTLV-1感染細胞株で、MGMT遺伝子の発現が検出されないことが見い出された。MGMTは、変異原性O6-メチルグアニンからメチル基を除去するDNA修復酵素である。従って本論分の知見はHTLV-1感染細胞ではアルキル化反応によって生じるDNA損傷を修復することができないことを意味する。次に、この発現低下のメカニズムを明らかにする目的で、ゲノムの構造異常による不活性化、プロモーター活性の低下、mRNAの不安定化、について検討したが、いずれも正しくないことが示された。 次にMGMT遺伝子の発現低下をもたらすHTLV-1遺伝子を同定する目的で、HTLV-1各遺伝子を発現する細胞株を作成し解析した結果、Taxを発現するクローンの半分にMGMT遺伝子発現の低下が見られた。さらに、発現低下が見られたTax発現クローンをサブクローニングしたところ、サブクローンの約半分がMGMTを全く発現していなかった。このことから、少なくともHTLV-1遺伝子産物TaxによってMGMT遺伝子発現の抑制が引き起こされることが示された。しかし、Taxの発現下でもMGMTの発現が高レベルで存続するクローンもあり、その効果の一過性が推定された。 一方で、MGMT遺伝子の高メチル化がその高い発現と相関することが報告されている。そこで、TaxによってMGMTの発現が低下した細胞クローンのDNAのメチル状態を解析した結果、MGMT高発現の細胞では高メチル化、MGMTの発現がない細胞株では低メチル化状態にあることが明らかとなった。そこで、HOS細胞をDNAメチルトランスフェラーゼの阻害剤である5-aza-C(5-azacytidine)で処理し、MGMT遺伝子発現を検討したところ、MGMT mRNA量は著しく減少することがわかった。以上の結果から、MGMT遺伝子の低メチル化がMGMT遺伝子発現抑制の原因になりうることが示された。 以上、本研究においてほとんど全てのHTLV-1感染細胞においてMGMT遺伝子発現が検出されないこと、HTLV-1遺伝子産物Taxがこの現象を引き起こすこと、MGMT遺伝子発現とMGMT遺伝子のメチル化状態との間に相関性があり、MGMT遺伝子の低メチル化がMGMT遺伝子発現抑制の原因になりうることを明らかにした。このようにこの研究はHTLV-1による発がん機構に、今まで知られていない新しい視点を与えるもので、ウイルス腫瘍学や分子生物学の進展に寄与するものであり、博士(薬学)の学位に相当すると判断される。 |  、granulocyte/macrophage colony-stimulating factor、c-Fos、c-Junなどの細胞増殖に関わる遺伝子発現を活性化する。また、Taxは細胞をトランスフォームさせる能力を持つ。しかし、Taxの直接作用だけではATL発症に充分ではないことが示唆され、HTLV-1感染後に細胞の遺伝子発現の変化が必要であることが想定されている。
、granulocyte/macrophage colony-stimulating factor、c-Fos、c-Junなどの細胞増殖に関わる遺伝子発現を活性化する。また、Taxは細胞をトランスフォームさせる能力を持つ。しかし、Taxの直接作用だけではATL発症に充分ではないことが示唆され、HTLV-1感染後に細胞の遺伝子発現の変化が必要であることが想定されている。