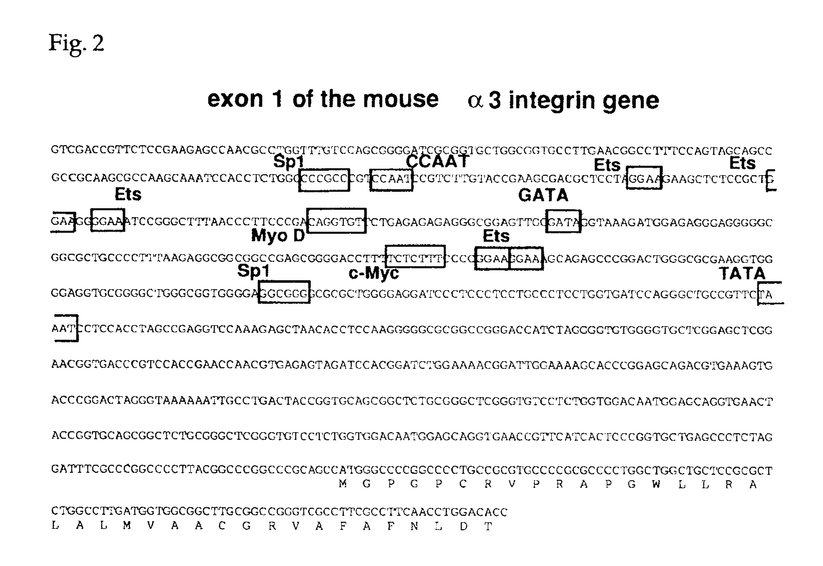
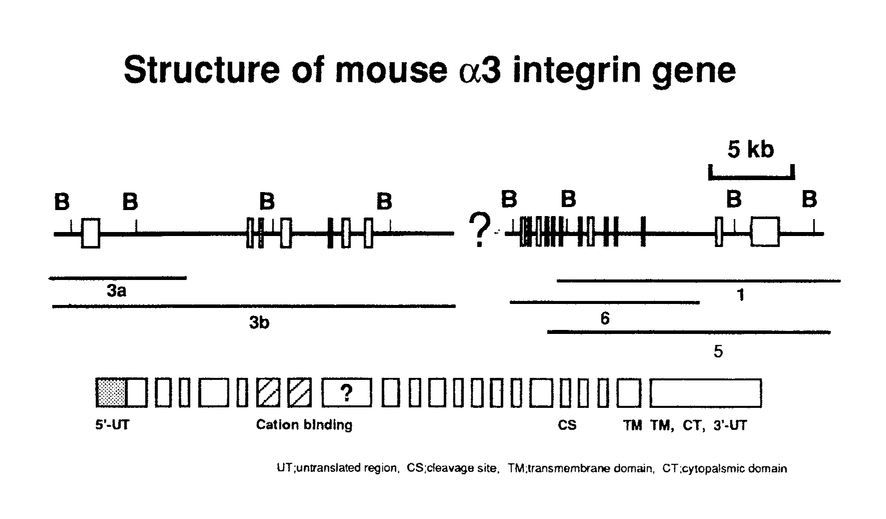
本論文は、インテグリンファミリーに属する接着分子のひとつで、比較的その機能の解析が進んでいない、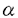 3 3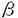 1インテグリン(VLA-3)の3サブユニットの、ゲノムDNAの構造とプロモーター活性を解析した結果をまとめたものである。 1インテグリン(VLA-3)の3サブユニットの、ゲノムDNAの構造とプロモーター活性を解析した結果をまとめたものである。 第一章はこの研究の背景を述べている。まず、多細胞生物の生物学における細胞接着分子、とりわけインテグリンの重要性を手短にまとめ、なぜ3インテグリンの遺伝子構造及びプロモーターの解析が緊急の課題と考えられるかが述べられている。さらに、インテグリン全般に関してこれまでに得られている知見が簡潔にまとめられている。 第二章では、マウス3インテグリンのプロモーターの解析結果が述べられている。マウスゲノムDNAライブラリーを、3インテグリン全長cDNAをプローブにしてスクリーニングした結果、5つの陽性クローンが得られた。制限酵素を用いた解析およびサザンブロッティング解析の結果、2つのクローンが3インテグリンのN末端領域をコードしていることが判明した。塩基配列を解析した結果そのうちのひとつのクローンは約4kbの5’上流領域と、第1エクソンと第1イントロンの一部を含んでいることが判明した。プライマー伸長分析の結果、主な転写開始部位が翻訳開始部位の328bp上流に位置し、そのすぐ上流にはTATAボックス様の配列が存在することが判明した。そこで学位申請者は3インテグリンの転写制御機構を解明するため、5’上流領域4kbをCATレポータープラスミドに挿入した発現ベクターを用いてプロモーター活性を検討した。線維芽肉腫細胞と乳癌細胞の細胞株においてプロモーター活性が認められた。この5’上流部分から一部の配列を削除して、CATアッセイによりプロモーター活性を比較検討した結果、最下流の0.5kbの断片が最も強いプロモーター活性を示すことが判明した。この部分の塩基配列を解析した結果、CAATボックスやGCアイランド、GATAモチーフ、またEts、Myo-D、c-Myc等の産物が結合しうる配列を見いだした。以上のように3インテグリンの発現制御を解明するための基盤を確立することができた。 第三章は、5つのゲノムクローンの配列解析結果に基づいて、全配列の推定を試みた結果が述べられている。配列が決定したのは、全体の87%で、518bpに相当する部分はゲノムクローンが得られていないことが判明した。全体は、30kb以上で、20以上のエクソンから成ることが明かとなった。すべてのイントロンーエクソン結合部位は、mRNAスプライシングの規則配列を保持していた。また、第1エクソンには、5’非翻訳領域とシグナルペプチドと3インテグリンポリペプチドのN末端37アミノ酸がコードされていた。このようなアレンジは、既に知られていた4および5インテグリン遺伝子のゲノム配列と類似しているが、2インテグリンとは異なることが判明した。3カ所存在する推定上の2価陽イオン結合ドメインや膜貫通ドメインは、複数のエクソンにまたがっていることが判明した。3インテグリンには細胞内ドメインの異なる2つのバリアントが存在し、これらはマウスにおいて臓器特異的に発現することが知られていたが、今回ゲノムDNAを解析した結果この2種の3インテグリンがmRNAスプライシングの違いで生じることが証明された。これらの結果はまた、インテグリンファミリーに属する接着分子の進化を理解する上でも、欠くことのできない情報を与えたと言える。 以上、学位申請者韓善愛は、マウス3インテグリン遺伝子をクローニングして、プロモーター領域とエクソンーイントロン構造を解析した。ゲノム遺伝子の一部分にクローニングが完成していないところがあるものの、この研究は、多細胞生物における細胞相互作用を規定している重要な接着分子である3インテグリン遺伝子の、細胞分化に伴う発現調節を理解する上で極めて有意義である。従って、これらの研究成果は細胞生物学及び腫瘍生物学に資するところが大であり、学位申請者は博士(薬学)の学位を受けるに十分であると判断した。 |  3インテグリン遺伝子の特徴
3インテグリン遺伝子の特徴 EMBL3で作製したマウスゲノムDNAライブラリー(1×106pfu)を、
EMBL3で作製したマウスゲノムDNAライブラリー(1×106pfu)を、