本研究は、神経発生過程におけるイオンチャネルの発現制御のメカニズムを明らかにする目的で、ホヤの分割抑止割球の系を用いて、この系でNa+チャネルをコードすると思われるcDNAクローンをえて、これをTuNaIとなづけ、このTuNaIと実際の電流との対応付けを行ったものであり、次の結果を得ている。 1、マボヤ幼生のcDNAライブラリーより、PCR法およびスクリーニング法を用いて他の動物のNa+チャネルに相同の構造をもつクローンを得、TuNaIとなづけた。 2、マボヤの正常な発生過程において、どの段階でTuNaIが発現されるかをNorthern blot法およびRT-PCR法を用いて調べたところ、中期から後期胞胚期にかけて転写が始まることが明らかとなった。この転写パターンからTuNaIは3種のNa+チャネルのうちtype-C電流に対応することが示唆された。 3、TuNaIが神経特異的に発現しているかどうかをみるため、幼生期の個体にTuNaIをプロープとしてin-situ hybridization法をおこなった。その結果、解剖学的に神経細胞の存在が確認されている部位に一致してTuNaIの発現が認められ、TuNaIが神経特異的に発現していることが確かめられた。 4、神経発生に特定の細胞間の接触が関与しているという仮説を単一細胞レベルで検証するため、分割抑止割球を用いた2細胞の神経誘導の系で、TuNaIをプロープとしてin-situ hybridizationをおこなった。その結果神経に分化する運命をもつa4-2割球は、A4-1割球と接触させた条件下で培養するとTuNaIを発現するが、単独で培養した場合や、A4-1以外の細胞、例えばb4-2割球と接触した状態で培養してもTuNaIは発現しないことがわかった。さらに正常のembryoからA4-1割球を除去したところ、残りの胚からTuNaIの発現は認められなかった。 5、a4-2割球は、神経誘導を受けると、分化の過程で電気生理学的に3種類のNa+電流を発現することがわかっている。TuNaIがこの3種類のうちのいずれに相当するかをみるためには、TuNaIをXenopus未受精卵などの再構成系で発現させて電気的な性質を調べることが一つの有効な方法と考えられたが、実際にTuNaIのmRNAをXenopus未受精卵に導入したところ、Na+電流の発現は認められなかった。そこで、神経誘導をうけたa4-2割球にTuNaIの翻訳開始部位にあたる配列に対するアンチセンス配列を注入し、電気生理学的にその影響を調べた。その結果、神経分化の最終段階で発現されるtype-C電流が特異的に抑制され、TuNaIはtype-C電流に相当するチャネルをコードしていると考えられた。 6、ホヤの割球の細胞分割を止めた系で、外来性にチャネル分子のmRNAを発現させることを目的として、まずコントロールとして、 -galactosidaseをHerpes Simplex Virusのthymidine kinase 5’非翻訳領域のcDNAの下流につないだDNAからin-vitroでRNAを合成し、cytochalasinBで細胞分割を止めた割球にmRNAを空気圧を用いて注入した。-galactosidaseの上流に核移行シグナルを入れたクローンを用いたため、RNAを注入した細胞では核が染まり、注入した-galactosidaseがタンパク質に翻訳されていることが確認できた。 -galactosidaseをHerpes Simplex Virusのthymidine kinase 5’非翻訳領域のcDNAの下流につないだDNAからin-vitroでRNAを合成し、cytochalasinBで細胞分割を止めた割球にmRNAを空気圧を用いて注入した。-galactosidaseの上流に核移行シグナルを入れたクローンを用いたため、RNAを注入した細胞では核が染まり、注入した-galactosidaseがタンパク質に翻訳されていることが確認できた。 7、スクリーニングおよびPCRで得られたcDNA断片をつなげて、TuNaIのcoding region全域と3’非翻訳領域を含む6.5kbpsの配列をもつプラスミドクローンを作った。クローン作成の途中でframe shiftなどのmutationが起こっていないことを確認するため、in vitro transcriptionおよびin vitro translationを行い、SDS-PAGEにて220kDa以上の長さのタンパク質が合成されることを確認した。先の-galactosidaseのコントロール実験の時に用いたHerpes Simplex Virusのthymidine kinase 5’非翻訳領域をTuNaIの配列の上流につなぎ、このプラスミドからin-vitroでRNAを合成し、神経に誘導した割球に注入した。その結果、RNAを注入していない割球で発現しているNa+電流に比べて、RNAを注入した割球では電流量の増加が認められ、外来性に導入したNa+チャネルが発現されているものと思われた。この際、電流の性質に大きな変化は見られず、type-C電流が増加していると考えられた。すなわちTuNaIはtype-C電流に相当するチャネルをコードしているという上記5の結論が支持された。 8、ホヤのNa+電流はtetrodotoxin(以下TTX)に対して感受性がない事が知られている。一方、TuNaIのporeを形成すると考えられる部位で、第1780アミノ酸はアラニンであるが、TTXに感受性を持つ他のNa+チャネルではこの部位はアスパラギン酸で、種を越えて保存されている。このアスパラギン酸を中性のアミノ酸に変異させるとTTXに対して非感受性になることがわかっている(Telau et al.,1991)。そこでTuNaIの第1780アミノ酸のアラニンをアスパラギン酸に変異させたクローンをつくり、薬理学的に内在性のNa+電流と区別することができるようにした。A1780Dを神経割球に発現させたところ、RNAを注入しない割球、またはコントロールのRNAを注入した割球では1 MのTTXで全く変化がみられないが、RNAを導入した割球では1MのTTXでNa+電流が減少した。減少した電流は不活性化が早く、type-Bの成分を含まず、type-C電流と考えられた。 MのTTXで全く変化がみられないが、RNAを導入した割球では1MのTTXでNa+電流が減少した。減少した電流は不活性化が早く、type-Bの成分を含まず、type-C電流と考えられた。 以上、ホヤ胚より、神経特異的に発現する、Na+チャネルの遺伝子TuNaIをクローニングし、実際に発現している電流との対応をアンチセンス法とmRNAの強制発現により解析した結果、nativeな発現系でNa+電流と遺伝子との関係が単一細胞のレベルで対応付けることが可能となった。本研究は神経細胞の分化過程におけるイオンチャネルの発現制御のメカニズムの解明に貢献を為すものと考えられ、学位論文として適当であると考えられる。 | 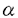 サブユニット以外のサブユニットによって制御されているという可能性も考えられる。実際、Xenopus未受精卵の再構成系で発現されたNa-チャネル
サブユニット以外のサブユニットによって制御されているという可能性も考えられる。実際、Xenopus未受精卵の再構成系で発現されたNa-チャネル