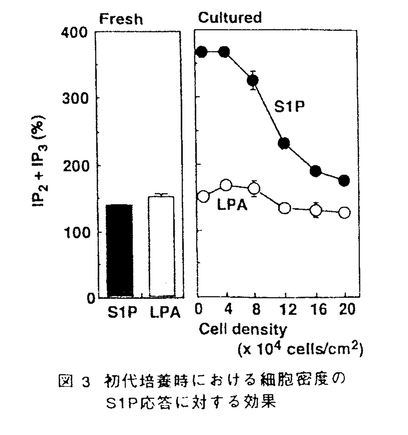
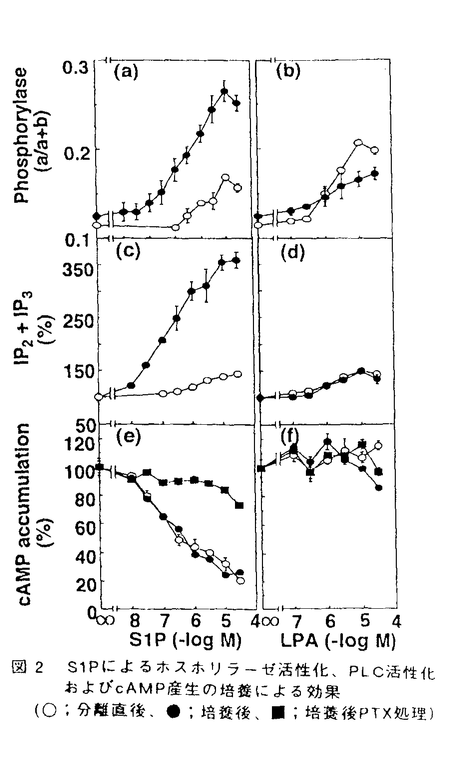
スフィンゴ脂質の代謝産物の一つであるスフィンゴシン1-リン酸(S1P)は、細胞の増殖や運動を調節することが知られているが、このS1Pは細胞内のCa2+貯蔵部位に直接作用してCa2+を放出させることから、細胞内シグナル分子として機能するものと考えられてきた。しかし最近になって、S1Pを細胞の外から加えても、細胞膜の受容体刺激を介するような細胞内初期応答、すなわち、百日咳毒素感受性のG蛋白質を介するホスホリパーゼC(PLC)の活性化、Ca2+動員、K+チャンネルの活性化、アデニル酸シクラーゼの抑制による細胞内cAMP生成の減少などが引き起こされるという知見が、種々の細胞において報告されている。これらの知見は、S1Pが細胞外及び細胞内の両方で機能する、極めてユニークな脂質性のシグナル分子である可能性を示唆している。 「ラット肝実質細胞におけるスフィンゴシン1-リン酸の作用とそのシグナル伝達機構」と題する本論文では、ラットの肝細胞を用いて、S1Pが肝細胞の重要な機能の1つであるグリコーゲン分解の律速酵素、グリコーゲンホスホリラーゼを活性化することを見い出し、そのシグナル伝達経路を解析している。また、その伝達経路が細胞周期に依存して変動することを、初代培養や再生肝などのモデル系で検討している。 スフィンゴシン1-リン酸(S1P)によるグリコーゲンホスホリラーゼの活性化とそのシグナル伝達経路の解析 コラゲナーゼ潅流法により調製したラット肝実質細胞をS1Pで刺激すると、PLCの活性化、イノシトール1,4,5-三リン酸(IP3)の産生、細胞内Ca2+濃度の上昇を経て、グリコーゲンホスホリラーゼが活性化された。このS1Pの作用は、細胞を百日咳毒素によって処理しても阻害されず、細胞を低密度で初代培養すると増強された。S1Pと構造が類似し、G蛋白質関連型受容体を介してその情報を伝達すると考えられているリゾホスファチジン酸に対する応答、あるいは、G蛋白質を直接活性化するNaFに対する応答は、初代培養による影響を受けなかった。したがって、肝細胞の初代培養によるS1Pのシグナル伝達の増強は、G蛋白質より上流の経路、おそらくは受容体の機能変化(S1P受容体数の増加または親和性の上昇)を介するもの推定された。 S1Pは一方でアデニル酸シクラーゼを抑制し、細胞内cAMP濃度を減少させたが、この作用は上記のPLC-Ca2+系の応答とは異なり、初代培養によっては影響を受けず、百日咳毒素処理によって完全に消失した。すなわち、S1Pがアデニル酸シクラーゼを阻害するシグナル伝達経路には、百日咳毒素に感受性のG蛋白質の介在が示唆された。また、初代培養による影響や百日咳毒素に対する感受性の差異から、S1PによるPLC-Ca2+応答系とアデニル酸シクラーゼ抑制系とは、異なった受容体-シグナル伝達系を介するものと推定された。 細胞外に添加したS1PのCa2+動員に関しては、2つの機構が提唱されている。すなわち、細胞内に入ってCa2+プールに直接作用する経路とPLCの活性化によるIP3産生を介する経路である。初代培養によってS1PによるPLC活性化、Ca2+動員、ホスホリラーゼ活性化のいずれもが増強されたことは、肝細胞においてはPLC-Ca2+応答系が重要であることを示唆している。 細胞周期に依存したS1PによるPLC-Ca2+応答系の増強 初代培養時におけるS1PのPLC-Ca2+応答系の増強は、肝細胞が増殖期に入るような低細胞密度培養下で顕著に観察された。肝細胞は細胞密度を高い状態に変えることによって、培養状態にもかかわらず増殖期への移行が抑えられるが、この時、S1PのPLC-Ca2+応答系の増強も抑えられた。さらに、肝臓の部分(約2/3を)切除により作成した肝再生状態のラット個体から分離された細胞では、S1Pに対するPLC-Ca2+応答性が正常肝細胞に比べて約2倍に増強することが確認された。すなわち、S1PによるPLC-Ca2+応答系の増強は肝細胞の細胞周期に依存し、その増殖期で顕著に認められた。 以上を要するに、本論文はラットの正常肝細胞において、その生理作用や作用機構が不明であったS1Pの作用を検討し、糖代謝への作用とそのシグナル伝達経路を明らかにし、脂質性の生理活性物質としてのS1Pの新たな生理的役割を見い出した。S1Pが2つの異なる受容体-シグナル伝達系を介して生理応答を惹起するという可能性の提示は、S1Pによる細胞応答の多様性を説明し、今後のS1Pシグナル伝達系研究の手がかりを提供している。また、細胞周期との関連性の検討は肝細胞にとどまらず、S1P-受容応答系(PLC-Ca2+系)の細胞生理学における役割の解明に大きな手がかりを提供しており、博士(薬学)の学位論文として十分な価値があるものと認められる。 | 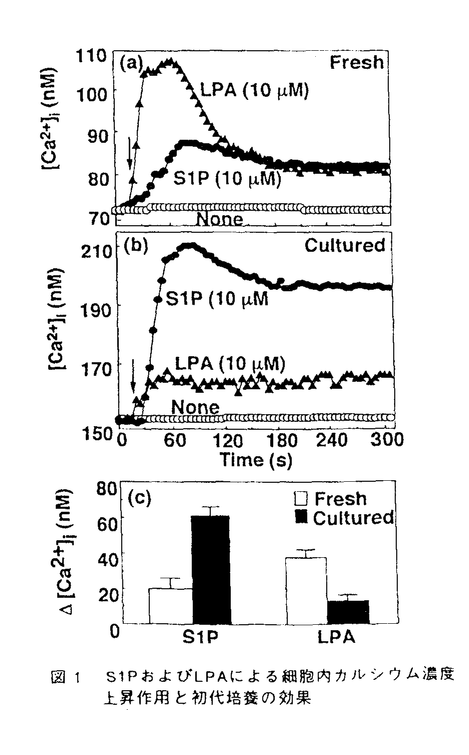
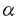 受容体アゴニスト、バソプレッシンなどのCa2+動員アゴニストによる細胞応答が減弱することが知られている。そこで24時間初代培養後の肝細胞におけるS1P、LPA応答を調べた。予想どおりLPAによるCa2+応答は減弱していたが、意外にも、S1Pによる応答は初代培養の前に比べてむしろ大きくなった(図1b)。ピーク値を比べてみると、LPAのCa2+応答が約半分くらい減少したのに対してS1PのCa2+応答は初代培養後約3倍にも増加していた(図1c)。
受容体アゴニスト、バソプレッシンなどのCa2+動員アゴニストによる細胞応答が減弱することが知られている。そこで24時間初代培養後の肝細胞におけるS1P、LPA応答を調べた。予想どおりLPAによるCa2+応答は減弱していたが、意外にも、S1Pによる応答は初代培養の前に比べてむしろ大きくなった(図1b)。ピーク値を比べてみると、LPAのCa2+応答が約半分くらい減少したのに対してS1PのCa2+応答は初代培養後約3倍にも増加していた(図1c)。