哺乳類の着床過程は、母体組織と胎仔組織との複雑な相互作用のもとに進行する極めてユニークな器官形成過程であるが、その分子・細胞機構は現在でも十分に解明されておらず、基礎・応用生物学における重要な課題の一つである。応用生命科学の分野では、特に新しい胚操作技術の開発、体外受精や胚移植の成功率の向上、不妊症の治療などの観点から、その機構解明が望まれている。 着床過程は、おおまかに、1)初期接着期、2)後期接着期、3)浸潤期に分けることができるが、その本質は、先に述べたように、母体と胎仔という遺伝的に異なる細胞間の複雑な相互作用であり、そのような相互作用を可能にしているのが、特異な分化を遂げた胚細胞、すなわち、栄養芽細胞である。着床過程に含まれるさまざまな現象の中でも、浸潤過程は最も複雑な過程の一つであり、従来、最も研究が遅れている分野の一つである。 近年、癌細胞による組織浸潤や転移と、着床期胚盤胞の栄養芽細胞による内膜浸潤が、機構の上で類似していることが推測されるところから、栄養芽細胞の浸潤機構の解明が、新しい癌治療法開発の手がかりをもたらすのではないかと考えられ、栄養芽細胞に対する関心が高まっている。 本論文において、我々は主として、マウス胚盤胞のin vitro培養系を用い、栄養芽細胞の分化を経時的に解析し、着床期栄養芽細胞の巨細胞化と巨大栄養芽細胞による組織浸潤の細胞機構について研究を進めた。 我々はまず、マウス着床期栄養芽細胞の巨細胞化に着目し、巨細胞化機構におけるNa+/H+exchanger系の関与について実験的解析を試みた。 一般に、細胞の体積変化には、細胞膜に存在するナトリウムイオン(Na+)と水素イオン(H+)の交換輸送系(Na+/H+ exchanger)が重要な役割を担っていることが知られている。しかし、栄養芽細胞の巨細胞化については、極めて著しい細胞の体積変化が起こるにも関わらず、Na+/H+ exchanger関与の可能性はもとより、その細胞機構について、従来の知見がほとんどない。そこで我々は、Na+/H+ exchangerに対する阻害剤であるアミロライド及びそのの誘導体で、より特異的な阻害剤であるHMA [3-amino-6-chloro-N-(diaminomethylene)-5-(1-homopiperidyl)-pyrazine-carboxamide]を用い、培養マウス胚盤胞に対する影響を調べた。マウス胚盤胞のin vitro培養系においてHMAは、trophoblast spreadingを濃度依存的に抑制する一方、栄養芽細胞の細胞増殖には影響を与えず、巨細胞化を特異的に抑制することを明らかにした。また、高濃度(15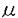 M)のHMAの存在下では、培養マウス胚盤胞の栄養芽細胞のtrophoblast spreadingは、非常に強く抑制されるが、内部細胞塊(ICM)においては、原始外胚葉への分化が認められ、HMAにより栄養芽細胞の分化が特異的に抑制されることを示した。以上の結果から、栄養芽細胞の巨細胞化にNa+/H+ exchangerが重要な役割を担っていることが推論された。 M)のHMAの存在下では、培養マウス胚盤胞の栄養芽細胞のtrophoblast spreadingは、非常に強く抑制されるが、内部細胞塊(ICM)においては、原始外胚葉への分化が認められ、HMAにより栄養芽細胞の分化が特異的に抑制されることを示した。以上の結果から、栄養芽細胞の巨細胞化にNa+/H+ exchangerが重要な役割を担っていることが推論された。 さらに我々は、Na+/H+ exchangerに対する抗体を用い、培養胚盤胞におけるNa+/H+ exchangerの局在を蛍光抗体法により解析し、Na+/H+ exchangerが、培養胚盤胞のTrophoblast spreadingの辺縁部に局在することを明らかにした。一方、マウス7.5日胚の外胎盤錘の細胞を外科手術的に分離し、取り出して培養を行った場合についても、抗Na+/H+ exchanger抗体によりtrophoblastic outgrowthの辺縁部に位置する巨大栄養芽細胞にNa+/H+ exchangerの発現が特異的に認められた。以上の結果から、従来、全く知見の得られていなかった栄養芽細胞の巨大栄養芽細胞への分化の細胞機構にNa+/H+ exchanger系が関与している事が、結論づけられた。 次に我々は、栄養芽細胞の浸潤性に注目し、癌細胞による組織浸潤において中心的役割を果すことが知られているマトリックスメタロプロテアーゼ(MMP),特に、それらの中でも細胞膜貫通領域を有する膜型MMP(MT-MMP-1)の発現と、栄養芽細胞による組織浸潤との関連について研究を行った。 MMPファミリーは、よく保存されたドメイン構造をもち、近年、癌細胞による組織への浸潤過程の分子機構解明の見地から、多くのMMPファミリーが同定され、その機能解析が進められている。中でも、1994年に金沢大学の清木教授らのグループにより、新たに発見されたMT-MMP-1は、他の分泌型のMMPファミリーとは異なり細胞膜貫通ドメインを有するという著しい特徴を持っている。MT-MMP-1は、浸潤過程にある癌細胞の表面に発現し、そのエンドペプチターゼ活性により、他のMMP、MMP-2(ゼラチナーゼA)のプロエンザイムの活性化を部位特異的に行っているらしいことが示されている。すなわちMT-MMP-1の発現制御が、浸潤過程におけるMMP活性の調節機構の中心的な役割を果たしていることが推測される。 我々は、マウス栄養芽細胞による子宮内膜浸潤において、MMPの部位特異的活性制御に中心的役割を果たしていると推測されるMT-MMP-1の発現制御機構ついて研究を行い、以下のような結果を得た。 まず、RT-PCR法により、マウス初期胚におけるMT-MMP-1mRNAの発現レベルの解析を行ったところ、その発現量は、胚盤胞期に著しく増加することが明らかになった。また一方、培養胚盤胞において、MT-MMP-1mRNAの発現量は、培養が進むにつれ増加する傾向が認められた。さらに、交尾後7.5日齢のマウス胚の外胎盤錘の細胞について解析を行ったところ、MT-MMP-1mRNAの強い発現が認められた。この様なMT-MMP-1の発現様式は、マウス着床過程の初期浸潤期に、MT-MMP-1が、関与していること強くを示唆している。 さらに、抗MT-MMP-1抗体を用い、マウス培養胚盤胞および妊娠8日目の外胎盤錘の細胞培養において蛍光抗体法による解析を行ったところ、いずれの場合においても中心部において密に接した栄養芽細胞ではMT-MMP-1の発現は認められなかったが、outgrowthの辺縁部に位置する巨大栄養芽細胞においてMT-MMP-1の局在的な発現が認められた。この結果は、胚盤胞及び外胎盤錘の培養系において、活発にoutgrowthした栄養芽細胞の辺縁部の、おそらく、強い浸潤性を示すと予想される巨大栄養芽細胞が、特異的にMT-MMP-1を発現していることを示している。 他方で、我々は、培養胚盤胞において、MT-MMP-1の発現と局在が、白血病阻害因子(LIF)により制御されている可能性を示唆する結果を得た。すなわち、培養胚盤胞において、LIFにより、栄養芽細胞のoutgrowthの辺縁部でMT-MMP-1の発現している細胞数が増加する傾向が認められた。LIFは、マウスの着床成立に不可欠な因子として知られており、着床成立過程において、LIFによるMT-MMP-1や他のMMPの発現制御が重要な役割を果たすことが考えられる。 さらに我々は、培養胚盤胞及び外胎盤錘のoutgrowthした栄養芽細胞の辺縁部におけるMT-MMP-1の発現の局在に着目し、MT-MMP-1の発現制御機構に、細胞間相互作用が関与している可能性を検討する目的で研究を進めた。まず複数のマウス胚盤胞を、接近させて培養すると、栄養芽細胞のoutgrowthの辺縁部に位置した細胞であっても、隣接した胚盤胞のoutgrowthが接近し、互いにcontactした栄養芽細胞では、MT-MMP-1の発現は、失われることが確かめられた。 さらに、cell-cell contactによるMT-MMP-1の発現制御機構に関し実験的な解析を進める目的で、培養マウス乳腺上皮細胞株(HC11)を用いて実験を行い、非常に興味深い知見を得ることができた。すなわち、細胞どうしがcontactしていない増殖期のHC11細胞では、抗MT-MMP-1抗体による免疫染色により、MT-MMP-1の発現が認められたが、細胞どうしがcontactした状態の細胞では、MT-MMP-1の発現は、認められなかった。さらに、confluenceの状態のHC11細胞では、免疫染色によって検出可能なMT-MMP-1の発現は、消失していた。ノーザンブロット解析により、MT-MMP-1mRNAの発現量を調べた結果、MT-MMP-1mRNAの発現量は、増殖期からconfluenceの1日目まで変化せず、confluenceの2日目頃から、発現量が減少する傾向が見られた。この結果から、HC11においてMT-MMP-1の発現は、cell-cell contactによりdown-regulateされ、その発現調節は、おそらく翻訳レベルで制御されていると推論された。 MMPを介した細胞外マトリックスの分解、再構築は、動物の発生における組織構築に、重要な役割を果たすことが示唆されている。本論文で明らかにした、MT-MMP-1のcell-cell contactによる発現調節機構は、胎盤形成過程での栄養芽細胞による組織浸潤だけでなく、発生学における大きな課題である形態形成においても、その機構解明に重要な手がかりを与えるものと考えられる。また。癌細胞による組織浸潤機構の解明の見地からも意義深いものと思われる。 |