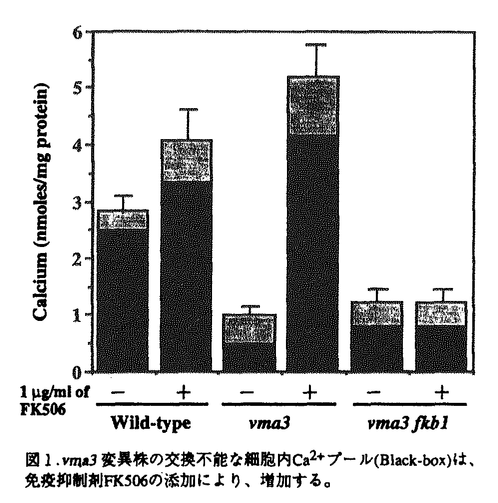
細胞質内の遊離Ca2+濃度(Ca2+i)を至適の低濃度領域(100-200nM)に維持することは,細胞機能の発現・調節に必須である。酵母細胞において、液胞は液胞膜H+-ATPaseによって形成されるプロトン駆動力を利用して、細胞内の95%以上のCa2+を蓄積しており、原形質膜と共に(Ca2+i)調節に重要な役割を果たしている。液胞膜H+-ATPaseの活性に欠損を持つvma変異株は、対数増殖期における(Ca2+i)が野生株(150nM)に比べ約6倍(900nM)に上昇している。このことは(Ca2+i)調節における液胞膜H+-ATPaseの役割の重要性を示唆している。論文提出者,谷田は,vma変異株の多面的表現型、特にCa2+感受性に注目して(Ca2+i)調節に関わる新たな因子を解明するため,下記の分子生物学的・分子生理学的研究を行った。 1)vma変異株が免疫抑制剤FK506(1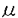 g/ml)およびサイクロスポリンA(50g/ml)に感受性を示すことを発見した。FK506とサイクロスポリンAは、それぞれの細胞質内結合蛋白質FKBP-12(Fkb1p)およびサイクロフィリンA(Cyp1p)と複合体を形成し、カルシニューリンの活性を阻害する。それぞれの結合蛋白質の遺伝子破壊変異(fkb1変異及びcyp1変異)導入実験によって、FK506/FKBP-12複合体及びサイクロスポリンA/サイクロフィリンA複合体がカルシニューリンの活性を阻害し、その結果としてvma3変異株の増殖を阻害する事を示した。また、vma3変異株にカルシニューリンの活性変異(cnb1変異)を導入したところ、合成致死を示した。このことにより、カルシニューリンの新しい機能はvma変異のもとに容易に検出できることを明らかにした。 g/ml)およびサイクロスポリンA(50g/ml)に感受性を示すことを発見した。FK506とサイクロスポリンAは、それぞれの細胞質内結合蛋白質FKBP-12(Fkb1p)およびサイクロフィリンA(Cyp1p)と複合体を形成し、カルシニューリンの活性を阻害する。それぞれの結合蛋白質の遺伝子破壊変異(fkb1変異及びcyp1変異)導入実験によって、FK506/FKBP-12複合体及びサイクロスポリンA/サイクロフィリンA複合体がカルシニューリンの活性を阻害し、その結果としてvma3変異株の増殖を阻害する事を示した。また、vma3変異株にカルシニューリンの活性変異(cnb1変異)を導入したところ、合成致死を示した。このことにより、カルシニューリンの新しい機能はvma変異のもとに容易に検出できることを明らかにした。 2)vma3変異株のCa2+感受性に対するFK506の効果を詳しく調べ、薬剤の添加量に応じてその感受性が上昇することを認めた。このことは,FK506によってカルシニューリンの活性が阻害されることにより、(Ca2+i)に異常が生じたと考えられる。このことを検証するため,FK506存在下でのvma3変異株の(Ca2+i)をFura-2/顕微測光法により測定した。その結果、FK506の添加によりvma3変異株の(Ca2+i)が減少することがわかった。そこでvma3変異株の細胞内Ca2+プールの変化を45Caを用いて調べた。細胞内Ca2+プールは、(Ca2+i)を反映する短時間で外界のCa2+と交換可能な遊離Ca2+プールと細胞内小器官に取り込まれているために短時間では交換不能な貯蔵Ca2+プールからなる。vma3変異株においては細胞内の主要な貯蔵Ca2+プールである液胞へのCa2+の取り込みが起こらないために、交換不能な貯蔵Ca2+プールが野生株に比べて20%程度に減少している。ところがFK506を添加すると、この貯蔵Ca2+プールが約8.9倍に増加し、この増加はFKBP-12を欠損させたvma3 fkb1二重変異株では認められなかった。このとき液胞の酸性化は認められなかったことから、Ca2+が液胞に蓄積された可能性は低い。また、野生株においても、FK506添加による交換不能な貯蔵Ca2+プールの増加が認められた。野生株にFK506を添加した場合及びcnb1変異を導入した場合には、野生株はより高濃度のCa2+に対して耐性を示した。以上の結果から、FK506によってカルシニューリン活性が阻害されると、液胞以外の細胞内小器官へのCa2+の流入が起こることが示唆された。 3)vma3変異株においては、交換不能な貯蔵Ca2+プールが20%程度にまで減少している事を利用して、CLS2遺伝子産物が通常の培養条件において、細胞内貯蔵Ca2+プールの形成に関与していることを示した。また、cls2 vma3二重変異株が免疫抑制剤FK506に対して、vma3変異株の10倍以上も感受性であることを発見し、FK506を培地に添加した際、cls2 vma3二重変異株の交換不能な貯蔵Ca2+プールが相乗的に増加することを示した。CLS2遺伝子産物は小胞体に局在することから、CLS2遺伝子産物が小胞体膜上で細胞質内遊離Ca2+濃度調節に関わっている可能性を示唆した。 4)液胞膜H+-ATPaseと共に(Ca2+i)調節に関わる因子を探索するために、vma3変異株の多面的表現型に対する抑圧変異の単離を行った。得られた115株の劣性抑圧変異株は5つの相補群に分かれ、この抑圧変異をsuv変異と命名した。suv1変異、suv2変異、suv3変異、suv4変異はvma3変異株のCa2+感受性、中性pH感受性、Pet-表現型を抑圧した。一方、suv5変異は、Pet-表現型のみを抑圧した。すべてのsuv変異はvma3変異株の表現型の他にvma1変異株,vma2変異株,およびvma11変異株の表現型をも抑圧したので、suv変異は液胞膜H+-ATPaseの機能欠損を抑圧する新たな細胞質内Ca2+濃度維持・調節に関与する新規遺伝子の変異群であると結論した。 以上,論文提出者は,カルシニューリンによる新たな細胞質内遊離Ca2+濃度調節機構を解明し,さらに,(Ca2+i)の調節・維持にかかわる新しい遺伝子群(cls2,suv1-suv5)の発見に成功した.よって,審査委員は全員一致して,この業績は博士(理学)の学位にふさわしい成果であると認めた. |