骨形成を担う骨芽細胞の機能は多くの因子により調節されている。ホルモンとしては従来より知られている副甲状腺ホルモン(PTH)などの古典的Ca調節ホルモン以外にも性ホルモン、グルココルチコイドなどのステロイドホルモンが重要な役割を演じていること、さらにサイトカイン・成長因子の作用がホルモン以上に重要な役割を果たしていることが主に培養骨芽細胞を用いた近年の検討により明らかになってきた。成長因子は、骨細胞・あるいは近傍の細胞により産生・分泌されるだけでなく、骨基質中にも貯蔵されており、それらがホルモンなどの刺激に反応し活性化・不活化され、局所因子としてより直接的に骨芽細胞機能を調節しているものと考えられている。これらの成長因子の中でインスリン様成長因子-I(IGF-I)およびIGF-IIはtransforming growth factor-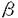 (TGF-)とならび骨形成促進因子として注目を浴びてきた。 (TGF-)とならび骨形成促進因子として注目を浴びてきた。 主にラット・マウス由来の培養骨芽細胞による検討から、培養骨芽細胞はIGFに対する受容体を有し、IGFに反応して増殖および主要基質蛋白であるI型コラーゲンの産生が促進されることが知られていた。ラットやマウス由来の骨芽細胞モデルはIGF-Iおよび-IIを産生する。特にラットの初代培養骨芽細胞(ROB)では、エストロゲンおよびPTHによる細胞の増殖やコラーゲン産生の促進が、抗IGF-I中和抗体で阻害されることから、IGF-Iについてはこれらのホルモンの骨形成促進作用を媒介するオートクリン物質としての役割が想定されていた。さらに、ヒトの血中IGF-I濃度が加齢により減少することから、骨形成の低下を主体とする老人性骨粗鬆症の発症機序にも関わっている可能性が考えられいた。 しかしながら、従来IGF-Iのオートクリン物質としての作用の検討は殆ど総て齧歯類由来の細胞を用いて行われており、ヒト由来の骨芽細胞はIGF-Iを産生しないとする報告をはじめ、ヒトでの骨形成調節因子としてのIGFの役割は明らかではなかった。一方、IGF-IIは、ヒト骨基質中に存在する成長因子としてTGF-と並んで最も多く含まれること、いくつかのヒト骨芽細胞が産生することが報告されていたが、ROBにおいてはIGF-Iと異なり骨代謝に関わるホルモンによりその産生が調節されておらずその生理作用については不明な点が多く残されていた。更に、本研究に着手した頃より、IGF作用を制御する高親和性結合蛋白、IGF結合蛋白(IGFBP)の存在が明らかにされ、培養骨芽細胞も種々のIGFBPを産生・分泌することが報告された。IGFBPには、IGFBP-1〜6の6種の存在が現在確認されているが、骨芽細胞モデルが分泌するIGFBPには、ラットとヒト由来の細胞との間に相違があるだけでなく、ヒト由来の細胞モデルの間でも、骨肉腫細胞株と正常海綿骨由来の初代骨芽細胞様細胞(hOB)との間には相違が認められることが報告された。IGFBPのIGFに対する親和性は、IGF受容体のそれに匹敵するため、IGF作用の検討にはIGFBPの変化を検討することが必須なだけでなく,従来抗IGF抗体を用いて行われてきたIGFのimmunoassayや中和実験の結果の解釈に問題があることが明らかとなってきた。 そこで、ヒトの骨形成におけるIGFの生理的意義を明らかにするためには、ヒト骨芽細胞モデルにおいてIGFBPを含むIGF系を構成する要素を個々に解析することが重要であると考え以下の検討を行った。 まず、疑問視されていたIGF-Iの産生を、hOBだけでなく、SV40 T antigenの導入により不死化したhOB細胞(HOBIT)および4種のヒト骨肉腫由来細胞株において、ribonuclease protection assay (RPA)およびreverse transcription-polymerase chain reaction(RT-PCR)を用いIGF-ImRNAを検出することにより比較検討した。その結果、hOBのみがIGF-I mRNAを発現していることが明らかとなった。一方、IGF-II mRNAはhOB以外にも3種類のモデル細胞で発現されていた。さらに、hOBの培養上清中にimmunoreactiveなIGF-Iおよび-IIが検出された。これは、ヒト骨芽細胞自身がIGF-IIだけでなく、IGF-Iも産生しうることを示す初めての結果であり、ヒトの骨芽細胞機能調節にもautocrine因子としてIGF-Iが作用する可能性を示す成績であった。また、hOB以外の細胞におけるIGF-I発現の欠如は、形質転換がIGF遺伝子の発現に影響を与える可能性を示唆するものと考えられた。従って、以下の検討はhOBを用いて行った。 IGF-Iは、alternative splicingにより、4種のmRNAを産生するが、骨においてはこれら種々のmRNAの存在様式は明らかではなかった。そこで、RPAおよびexon特異的なprimerを用いたRT-PCRを利用し、hOB細胞が総ての種類のIGF-ImRNAを発現し、これらのmRNAの相対的発現量が、他の組織と類似していることを明らかにした。次に、hOBが発現するIGFBPを検討し、hOBはIGFBP-2を除くすべて、すなわちIGFBP-1、およびIGFBP-3〜-6を発現していることを明らかにした。ROBでは、IGFBP-1の発現は認められず、hOBでは確認不能であったIGFBP-2は容易に検出されることが報告されていることから、IGFBP遺伝子の発現に種差のある可能性が明らかとなった。以上の検討およびaffinity cross-linkingの結果から、hOBはIGF-Iおよび-II、5種類のIGFBP、および2種類のIGF受容体を発現する系であることを明らかにできた。 次に、骨芽細胞機能に影響を与える他の因子がhOBのIGF系に変化をもたらすか否かを検討する目的で、ホルモンである糖質コルチコイド(GC)とサイトカインであるTGF-の効果を検討した。GCは、培養骨芽細胞では、分化を促進させると共に短期の投与では増殖も促進させる場合があることが報告されていた。hOB細胞において、IGF-Iは単独では殆ど増殖促進作用を示さなかったが、5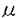 Mのcortisolによる48hの前処置は、それ自身で殆ど増殖効果を示さず、IGF-Iに対する増殖促進反応をもたらすことが明らかとなった。そこで、GCによるIGF-I反応性獲得にIGFBPが関わっている可能性を検討した。その結果、hOBの培養上清中にWestern ligand blot (WLB)により蛋白として検出可能なIGFBP-3、-4、および-5がいずれもGC添加により、時間・用量依存的に減少すること、hOB細胞のIGFBP-3、-4、および-5mRNAレベルも低下すること、更にhOBが産生するこれら3種のIGFBPに対する蛋白分解酵素活性には変化を与えないことを明らかにした。一方、GCはWLBによっては、培養上清中で検出不能であったIGFBP-6 mRNAレベルは変化させず、IGFBP-1 mRNAレベルは著明に増加させた。従って、GCはhOBが分泌する主要なIGFBPであるIGFBP-3、-4、および-5の細胞周囲濃度を主にその産生抑制により低下させること、またGCのIGFBPに対する作用は個々のIGFBPに特異的であることが明らかとなった。一般に、IGFBPは細胞外において蛋白分解酵素の修飾を受けないままの形で存在する場合、IGF作用を抑制することから、hOBにおけるGCによるIGF-I反応性獲得には、これら3種のIGFBPの産生低下が関わっている可能性が明らかとなった。 Mのcortisolによる48hの前処置は、それ自身で殆ど増殖効果を示さず、IGF-Iに対する増殖促進反応をもたらすことが明らかとなった。そこで、GCによるIGF-I反応性獲得にIGFBPが関わっている可能性を検討した。その結果、hOBの培養上清中にWestern ligand blot (WLB)により蛋白として検出可能なIGFBP-3、-4、および-5がいずれもGC添加により、時間・用量依存的に減少すること、hOB細胞のIGFBP-3、-4、および-5mRNAレベルも低下すること、更にhOBが産生するこれら3種のIGFBPに対する蛋白分解酵素活性には変化を与えないことを明らかにした。一方、GCはWLBによっては、培養上清中で検出不能であったIGFBP-6 mRNAレベルは変化させず、IGFBP-1 mRNAレベルは著明に増加させた。従って、GCはhOBが分泌する主要なIGFBPであるIGFBP-3、-4、および-5の細胞周囲濃度を主にその産生抑制により低下させること、またGCのIGFBPに対する作用は個々のIGFBPに特異的であることが明らかとなった。一般に、IGFBPは細胞外において蛋白分解酵素の修飾を受けないままの形で存在する場合、IGF作用を抑制することから、hOBにおけるGCによるIGF-I反応性獲得には、これら3種のIGFBPの産生低下が関わっている可能性が明らかとなった。 一方、TGF-は骨形成促進物質としてROB、hOBの両者において増殖およびコラーゲン合成を促進するが、ROBではIGF-Iおよび-IIの産生を抑制することが報告されていた。今回、TGF-がhOBでは強力にIGF-ImRNA発現を促進し、初代培養の骨芽細胞間にも種差が存在することが明らかとなった。一方、forskolinなど細胞内cAMPを上昇させる物質に対するIGF-ImRNAの反応にはROBと差がなく、hOBにおいてもIGF-ImRNAレベルの増加が認められた。hOBにおけるforskolinおよびTGF-によるIGF-ImRNA増加の時間経過は前者が3-6hをピークとし、24h後には効果が消失するのに対し、後者の作用は24-48h後をピークとする異なる時間経過を示した。そこで、これら2つの情報伝達系、作用の時間経過を異にする物質が、各クラスのIGF-I mRNA発現に及ぼす作用をRT-PCRにより検討した。その結果、forskolin、TGF-はいずれも総てのクラスのIGF-ImRNAレベルを増加させ、総てのクラスのmRNAがIGF-ImRNAの増加に関与していることが明らかとなった。TGF-は、hOB培養上清中のIGFBP-4レベルを主にIGFBP-4分解酵素の活性亢進を介して減少させることが報告されていたことから、TGF-はIGF-Iの産生の促進と相まってIGF作用を促進する可能性があるものと考えられた。 本研究により、ラットやマウスだけでなくヒトの骨芽細胞もIGF-Iおよび-IIを産生しうること、またその作用がGCおよびTGF-により受容体結合より上流で調節されうることを示すことができ、ヒトの骨形成においてもIGF作用がautocrine因子として他の骨代謝調節因子の影響下で作用している可能性を示すことができた。 |