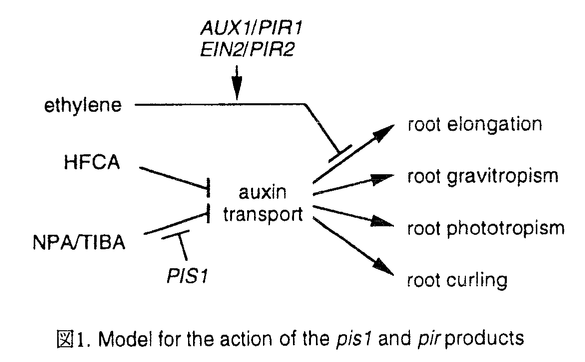
本論文は植物ホルモンの一つであるオーキシンの極性輸送という現象を、遺伝的な側面から新しい切り口で解析を試みたもので、序、第一章、第二章、考察の4部より構成されている。 序では、オーキシンの極性移動の研究の形態形成の研究における位置ずけ、研究の歴史と現状の分析を行ない、問題点の指摘と本研究の目的を設定している。その要約は下記のようなものである。 形態形成は、その最初の段階において極性が形成されることによって引起こされる。したがって、形態形成を理解するためには極性形成機構を理解することは重要である。このような観点から、様々な生物で極性形成機構の理解にむけての研究が盛んに行なわれている。植物にも種々の極性が存在するが、その中で特に有名なものの一つとしてオーキシンの極性輸送が知られている。この現象はオーキシンが重力の方向とは無関係に、常に植物の体軸の茎頂側から根端側へ輸送されるというものである。その原因としては、植物の体軸を構成する個々の細胞の細胞膜上にオーキシン輸送体(auxin-efflux carrier)が存在し、それが根端側により多く分布しているためと考えられているが、取り扱いの難しい膜タンパク質であり、その本体や分子機構はほとんど分かっていない。また、オーキシンの極性輸送は、NPA(N-1-naphthylphthalamic acid)、TIBA(2,3,5-triiodobenzoicacid)、HFCA(9-hydroxyfluorene-9-carboxylic acid)を代表とする化学構造の異なる物質群により特異的に阻害されることが知られており、それらの物質は総称してオーキシン(極性)輸送阻害剤と呼ばれている。今までに数多くの生理・生化学的な研究がこの輸送阻害剤を中心に行なわれてきたが、未だにその作用機構はほとんど明らかになっていない。そのような現状をふまえて、本論文提出者はオーキシン極性輸送系の分子機構の解明に遺伝的手法を適用することを試みた。すなわち、シロイヌナズナを用いて、オーキシン輸送阻害剤の一つであるNPAに対する感受性の変化した突然変異体の単離と解析を行なった。その結果は、第一章と第二章に記載されている。 [第一章の要約] 第一章には、NPA耐性突然変異体の結果が記されている。NPA耐性変異体は、EMS処理したM2種子より根の伸長阻害への効果を指標にして6個体単離された。これらは2つの遺伝子座に分かれそれぞれをpir1、pir2と名付けた。これらの突然変異体はNPAの根の伸長阻害に対しては耐性を示すものの、根の重力屈性阻害に対しての感受性は変化しなかったことから、NPAの効果のうち根の伸長阻害効果に特異的な突然変異体であると思われる。種々の植物ホルモンに対する応答を調べたところ、pir1変異体はオーキシンおよびエチレン生合成の前駆体であるACCに、pir2変異体はACCに明らかな耐性を示した。pir1変異体が根の重力屈性異常を示すことを考え併せ、両突然変異体は既に単離されている植物ホルモン応答の突然変異体の表現型と酷似していた。そこで相補性試験を行なったところ、pir1はaux1と、pir2はein2とそれぞれ遺伝子座が一致した。pir1突然変異体はオーキシン耐性突然変異体であり、pir2変異体はエチレン耐性突然変異体であることから、NPAの根の伸長阻害効果にこれら植物ホルモンが関与している可能性を強く示唆した。以下その点をさらに詳しく解析した。 エチレンの関与に関しては、野性型に対するNPAの伸長阻害効果が、エチレン作用の阻害剤である銀イオンやpir2エチレン非感受性変異によって部分的に回復したことから、NPAはエチレンを介して効果を示している可能性が考えられた。しかし、NPA投与によってエチレンの発生増加は認められず、組織内に存在するバックグランドレベルのエチレンがオーキシン輸送阻害剤の根の伸長阻害効果を強めていると考えられた。 オーキシン輸送阻害剤によるオーキシン輸送阻害と根の伸長阻害の関係はTIBAとそのアナログを用いて解析された。その結果、それら試薬によるオーキシン極性輸送の阻害と根の伸長阻害の間には相関関係は認められず、極性輸送阻害剤の根の伸長阻害には極性輸送を介さない阻害作用のあることが示唆された。 現在、オーキシン輸送阻害剤の根の伸長阻害効果は、オーキシン輸送阻害により蓄積した高濃度のオーキシンによって阻害効果が引起こされるという仮設によって説明されている。この点を含めNPAの根の伸長阻害効果におけるオーキシンの関与を二重突然変異体が用いて解析された。pir1変異体はオーキシンとエチレン双方に対し耐性を示すので、そのNPA耐性の性質はオーキシン耐性によるものか、エチレン耐性によるものかはっきりしない。しかし、pir2変異体はエチレンに対する感受性をまったく欠如しているが、オーキシンの感受性は正常な突然変異体であるので、pir2変異体とpir1 pir2変異体の両者を比較することにより、オーキシン耐性のみの影響を知ることができる。その結果、両者はNPAに対して同程度の耐性を示し、pir1変異体のNPA耐性がオーキシンに対する耐性ではなくエチレンに対する耐性によって付与されていることを示した。このことから、NPAの根の伸長阻害効果におけるオーキシンの寄与は認められず、従来の仮説は少なくともシロイヌナズナでは当てはまらないことを示している。 [第二章の要約] 第二章にはNPA高感受性突然変異体の単離とその変異体の性質について記載されている。高感受性変異体は耐性突然変異体と同様な方法でスクリーニングを行なった結果、2個体が得られたが、相補性試験の結果、同一遺伝子座(pis1と命名)の劣性突然変異体であることが明らかになった。 pis1突然変異体はオーキシン、エチレン、サイトカイニンなどの植物ホルモンや抗オーキシン剤に対する感受性の変化は認められず、上記耐性突然変異体(pir変異体)と異なり、植物ホルモンに対する感受性が変化したことが原因でNPAに対する感受性が変化したのではないと考えられた。また、NPAはシロイヌナズナ芽生えの成長阻害(ロゼットの直径および生重量)、根の伸長阻害、根の重力屈性阻害、根の光屈性阻害、根の回転成長阻害など非常に多岐にわたる効果を引起こすが、pis突然変異体はこれらの現象のすべてにおいて、約10〜30倍の高感受性を示した。NPAの幅広い効果に対して感受性が変化していることから、pis1突然変異体はオーキシン極性輸送系自体の変異体であることが強く示唆された。また、これらの屈性に関する結果から、一遺伝子座の変異により重力屈性と光屈性の反応が同時に異常になりうることがはじめて示された。このことは、重力屈性と光屈性の双方を調節する遺伝的因子が存在することを示し、またそれらの屈性反応にオーキシンの極性輪送が深く関わっていることを強く示唆するものでもある。さらに、pis1変異体のNPAに対する感受性の変化は、地下部の変化にくらべ極端に小さく、根に特異的な変異体と考えられた。このことはオーキシン極性輸送系が地上部と地下部では完全には一致しないことを示すものでもある。 一方、pis1突然変異体は化学構造の異なるオーキシン極性輸送阻害剤であるTIBAにたいしては、NPAと同様な高感受性を示したが、HFCAに対する反応は野生型に対する反応と変わりなかった。このことは、同じオーキシン極性輸送の阻害であってもその作用機作にはNPA/TIBAとHFCAの間で遺伝学的に分離される過程が存在することを示すものである。また、pis1変異はNPA/TIBAに特異的な作用点における変異であることを示している。pis1は劣性突然変異であり、loss-of-functionの変異であることが推測されるので、pis1遺伝子産物はNPA/TIBAがオーキシン輸送体を阻害する過程を負に調節していることが示唆される。また、pis1とpir1あるいはpir2との二重変異体を作成し、NPAに対する反応を調べた結果はpis遺伝子産物とpir遺伝子産物とはNPAの効果に対して独立に機能していることを示唆していた。このことは、pir突然変異体がエチレン非感受性の性質によってNPA耐性が付与されたとする第一章の結論と矛盾しない。 最後に考察では、従来オーキシン輸送阻害剤の作用は、輸送阻害剤によるオーキシン輪送の阻害によって蓄積した高濃度のオーキシンの作用として説明されてきたが、すべての現象にこの説明を適用することの危険性が指摘された。また、pis1に関しては極性輸送系自体に関わる新規の遺伝子であり、遺伝子を単離する必要性が述べられた。しかし、遺伝子座を決定したが周辺にマーカー遺伝子が少なく、その難しさと可能な方法について考察された。また、シロイヌナズナが遺伝的な解析にはすぐれているが、小型であり生化学的解析には必ずしも適当でないという実験材料上の制約から根のオーキシン輸送を測定できなかったが、その必要性と可能な方法についても考察された。 以上、本論文では、植物ホルモンのオーキシン極性輸送の解析に新たに遺伝的手法を導入することにより、多くの新知見を得、また、新しい道を切り拓いた。 よって本論文は博士(理学)の学位請求論文として合格と認められる。 |  M)に対しては若干の高感受性を、高濃度のNPA(10
M)に対しては若干の高感受性を、高濃度のNPA(10