学位論文要旨
| No | 112469 | |
| 著者(漢字) | 大木,理恵子 | |
| 著者(英字) | Ohki,Rieko | |
| 著者(カナ) | オオキ,リエコ | |
| 標題(和) | DNAの組み換えにおけるヌクレオソームの位相およびDNA湾曲部位の解析 | |
| 標題(洋) | Analysis of DNA recombination with respect to nucleosome phasing and DNA bending | |
| 報告番号 | 112469 | |
| 報告番号 | 甲12469 | |
| 学位授与日 | 1997.03.28 | |
| 学位種別 | 課程博士 | |
| 学位種類 | 博士(理学) | |
| 学位記番号 | 博理第3249号 | |
| 研究科 | 理学系研究科 | |
| 専攻 | 生物化学専攻 | |
| 論文審査委員 | ||
| 内容要旨 | DNAの組み換えは、大きくはゲノムの進化に伴う遺伝子の多様性獲得にはじまり、細胞の癌化や遺伝病に見られるゲノムの変化、免疫系の細胞で見られる分化の過程で厳密にコントロールされたV-(D)-J組み換えやクラススイッチング、DNAの複製及び修復に伴った組み換え、ウイルス感染などの外的要因によるDNAの挿入など様々な生物の機能に関わっている。in vivoにおける組み換えの機構は、人為的に構築したプローブを細胞に導入しその組み換え産物を解析したり、de novoの組み換え産物の解析を行うことによって研究が進められてきた。一方、組み換えに関与する蛋白質は数多く同定され、又組み換え部位の詳細な塩基配列の解析も行なわれたが、多くの場合組み換え部位に酵素の特異性を示すような特定の配列は発見されず、組み換えの全体像は未だ明かになっていない。近年、クロマチン構造と組み換えが関与していることを示唆するいくつかの報告があった。例えば、免疫グロブリン座位におけるV-(D)-J組み換えには、クロマチン構造の変化が必須であるというものなどがそれであるが、組み換えをクロマチン構造に重点をおいて調べている報告はあまりない。そこで、組み換えも他の生物学的事象と同様、クロマチン構造との関わりがあると考え、本論文では、ヒトHeLa細胞において多数存在することの知られるalphoid DNAが染色体外に切り出される際に生じる組み換え部位、さらにゲノムの組み換え体が安定に保持される例として白血病細胞に見られるc-mycと免疫グロブリン両座位間の組み換え部位の解析を行うことにより組み換えとクロマチン構造の関わりを、特にヌクレオソームの位相にポイントをおいて解析し、組み換えの機構の考察を行った。 染色体外環状DNAとは、核又は細胞質に存在する環状DNAであり細胞中の全DNAの0.001-0.1%存在し、繰り返し配列のものと同時に非常に低いコピー数のものも存在し、その多くは染色体由来であると考えられている。現在までに調べられた全ての組織や培養細胞で染色体外環状DNAの存在が認められており、そのサイズや量は細胞によりかなり異なるため、細胞の分化、増殖の状態等を反映するものと考えられている。又、これらは豊富に存在する自然の組み換え産物として組み換え部位の解析に適している。特にHeLa細胞由来の染色体外環状DNAには反復配列であるSau3A familyが濃縮して存在していることが知られており、染色体外環状DNAに対しSau3A familyをプローブとしたサザンハイブリダイゼーションを行うと特徴的な階段状のパターンが現われ、環状の組み換え体が多量に存在していることが伺える。 このようなSau3A familyの組み換え体の存在が確認された環状DNAより、選択的にDNA組み換え部位をクローニングし、解析を行った。計68の組み換え部位を解析したところ、4カ所(A-D)に集中することが判明した(図1)。B、Cはサブユニット間で特に相同性の高いところと一致していたが、A、Dは相同性の高さだけは説明できず、またこれらはヌクレオソーム構造をとったときにリンカーに隣接するコア領域に位置することより、組み換えにおいてヌクレオソーム構造が関与することが示唆された。 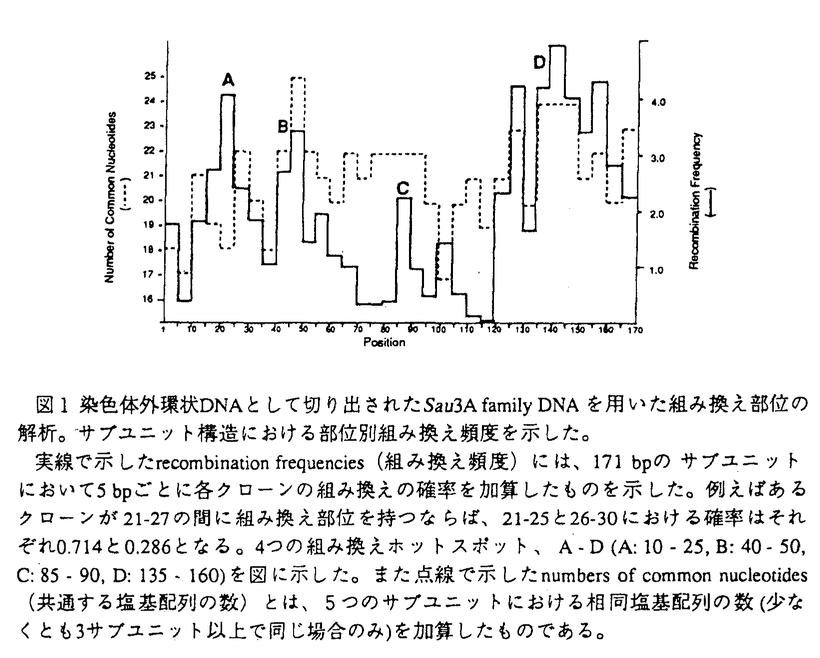 細胞が癌化した場合には、染色体DNAの組み換えが頻繁に起こることが知られている。例としては結腸癌にみられる、第5q及び第17p染色体の欠失やc-myc、neu、c-myb遺伝子等の増幅、胃癌にみられるK-sam遺伝子の増幅、乳癌におけるerbB様癌遺伝子ファミリーに属するHER-2/neuの増幅等がある。特に血球系の細胞においては、固形癌に比べ転座などの染色体異常の研究が進んでいる。例えばバーキットリンパ腫においては、その約90%が染色体8番と14番間の転座t(8;14)を起こし、残り10%はt(2;8)又はt(8;22)の転座を起こす。8番染色体側では常にc-myc遺伝子の存在する8q24の位置で転座が起きていること、2、14、22番染色体の転座部位には免疫グロブリン遺伝子が存在していることが知られている。また組み換え部位の塩基配列レベルの研究により転座に免疫グロブリンH鎖領域のクラススイッチ機構、あるいはV-(D)-J組み換え酵素が関与していると考えられている。このように組み換え部位についての解析が詳細に行われているc-myc、免疫グロブリン両座位間の転座を例に取り、alphoidの染色体外への切り出しの際に起こる組み換えに見られたような組み換えとクロマチン構造との関わりが、同様に存在するか解析を行なった。c-myc、免疫グロブリン両座位のヌクレオソーム位相を決定したところ、白血病培養細胞株Manca、BL67、ST486において両座位間の転座部位においてほぼ同じようなヌクレオソームの配置があることが判明し、やはり組み換えにヌクレオソーム構造が関わっていることが示唆された。さらにそれぞれの組み換え部位がヌクレオソーム構造内のどの部位に存在するかを調べたところ、alphoid DNAの組み換えで見られた組み換えのホットスポットの領域(ヌクレオソームとコアの境界近傍)とほぼ一致していた。このように組み換えにクロマチンが関与していることが示唆された。 以上のように組み換えにクロマチン構造が関与することが示唆されたが、それでは何によりクロマチン構造が決定されるかが次の興味の対象となる。染色体の高次構造であるクロマチン構造において折りたたみの第一段階はヌクレオソーム構造である。ヌクレオソームはコアヒストンの8量体にDNAが二巻きしたもので、ヌクレオソーム構造をとった時のDNAの構造は、ヌクレオソーム構造をとったときに占める位置により異なることが知られている。DNAの構造は遺伝子の転写や複製にも関与していると考えられることより、ヌクレオソームの位相がどのようにして決められているかは非常に興味深いことと思われる。ヌクレオソームの配置にはDNAの配列及び構造が関わっていることが知られ、特にDNAの折れ曲がり構造が存在するとヒストンへの親和性が高まると考えられる。木山らによりゲノム上に規則性を持ってDNAの湾曲部位が存在することが示され、この湾曲部位がヌクレオソームの配置に関与している可能性が示唆された。そこで、ヒト 湾曲部位のマッピングの結果、c-myc(全長8082bp)、免疫グロブリン(全長7427bp)両座位にも周期的に湾曲部位が存在しており、その間隔はそれぞれ693±282.3bpと653±180.8bpであった(図2)。従ってこの部位においても約680bpの周期性をもって湾曲部位が存在しており、これはヒト 本研究により以下のことが示された。 1.alphoid DNAの染色体外への切り出し、及び細胞の癌化に際して起こる転座における組み換えでは、ヌクレオソームの位相は保持される傾向がある。 2.DNAの組み換えは、DNAがヌクレオソーム構造を取ったとき、ヌクレオソーム構造上の特定の位置で高頻度に起きる。 3.ヌクレオソームの位相にはDNA湾曲部位が関与している。 DNAの組み換えにおいて、特にヌクレオソーム位相に着目して解析を行なった例は少なく、本研究は全く新しい方向性をもった研究と言える。 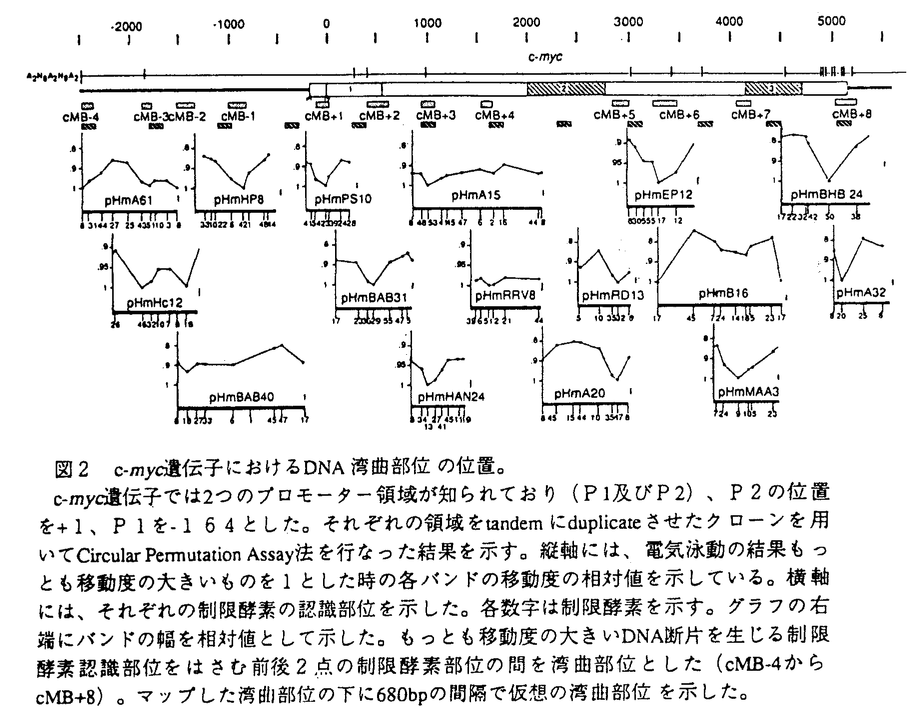 | |
| 審査要旨 | 本論文は、DNAの組み換えにおけるヌクレオソームの位相、およびDNA湾曲部位の影響について解析を行ったものである。DNAの組み換えは、遺伝子の進化、細胞の癌化、遺伝病、あるいは免疫系の細胞で見られるものなど様々な生物の機能に関わっている。本論文ではalphoid DNAが染色体外に環状のDNA(EC DNA)として切り出される際の組み換え部位と白血病細胞に見られるc-mycと免疫グロブリンの両遺伝子座間の組み換え部位の解析を行うことにより、組み換えとクロマチン構造、特にヌクレオソームの位相にポイントをおいて組み換え機構の考察を行った。 HeLa細胞のEC DNAには反復配列であるSau3A familyが濃縮して存在している。そこで、Sau3A familyが染色体外に切り出される際に生じるDNA組み換え部位を選択的にクローニングし、解析を行った。計68の組み換え部位を解析したところ、多くの組み換え部位が4カ所に集中していることが判明した。この内の特に大きなピークを示した2カ所はヌクレオソーム構造をとったときにコアとリンカーの境界に位置し、組み換えにおいてヌクレオソーム構造が関与することが示唆された。 第二点として、バーキットリンパ腫に見られる染色体8番と14番の間の転座部位の近傍のヌクレオソーム位相を決定し、組み換えとの関係について解析を行った。c-myc、免疫グロブリン両座位のヌクレオソーム位相を決定したところ、白血病培養細胞株において両座位間の転座部位においてほぼ同じようなヌクレオソームの配置があることが判明し、やはり組み換えにヌクレオソーム構造が関わっていることが示唆された。さらにそれぞれの組み換え部位がヌクレオソーム構造内のどの部位に存在するかを調べたところ、alphoid DNAの組み換えで見られた組み換えのホットスポットの領域(ヌクレオソームとコアの境界近傍)とほぼ一致していた。このように組み換えにクロマチンが関与していることが示唆された。 以上のように組み換えにクロマチン構造が関与することが示唆されたが、それでは何によりクロマチン構造が決定されるかが次の興味の対象となる。ヌクレオソームの配置にはDNAの配列及び構造が関わっていることが知られ、特にDNAの折れ曲がり構造が存在するとヒストンへの親和性が高まると考えられる。木山らによりゲノム上に規則性を持ってDNAの湾曲部位が存在することが示され、この湾曲部位がヌクレオソームの配置に関与している可能性が示唆された。そこで、ヒト 湾曲部位のマッピングの結果、c-myc(全長8082bp)、免疫グロブリン(全長7427bp)両座位にも周期的に湾曲部位が存在しており、その間隔はそれぞれ693±282.3bpと653±180.8bpであった。従ってこの部位においても約680bpの周期性をもって湾曲部位が存在しており、これはヒト なお、本論文は木山亮一氏、大石道夫氏と共同研究であるが、論文提出者が主体となって分析及び検証を行い、本論文に対し論文提出者の寄与が十分であると判断する。 よって、博士(理学)の学位を授与できると認める。 | |
| UTokyo Repositoryリンク | http://hdl.handle.net/2261/54566 |
 -globin座位に見られた湾曲部位の規則性が、c-myc、免疫グロブリンの両座位、さらには繰り返し配列であるSau3A family DNAに存在するかを調べ、ヌクレオソームの位相及び組み換えとの関わりについて考察した。
-globin座位に見られた湾曲部位の規則性が、c-myc、免疫グロブリンの両座位、さらには繰り返し配列であるSau3A family DNAに存在するかを調べ、ヌクレオソームの位相及び組み換えとの関わりについて考察した。