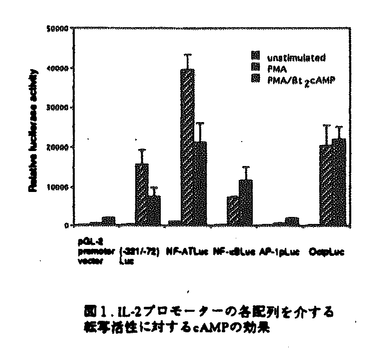
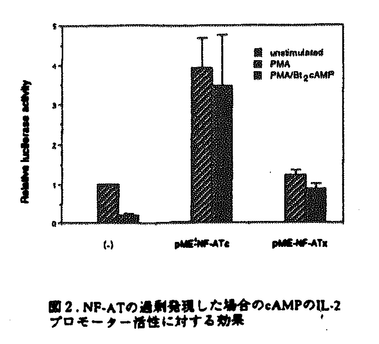
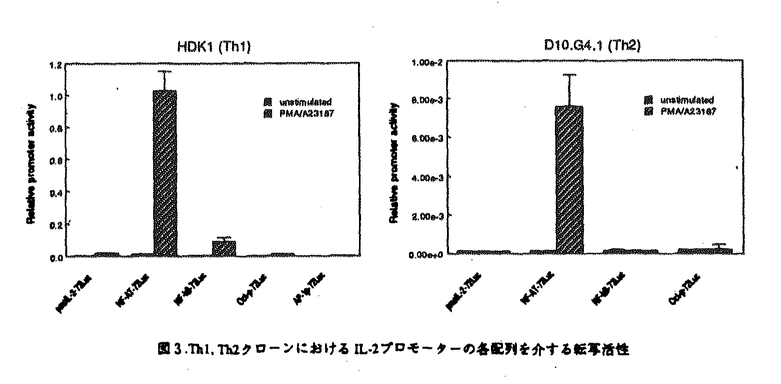
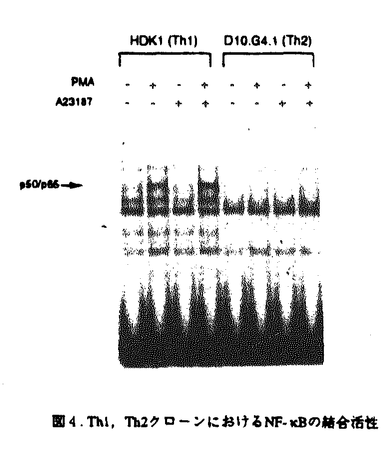
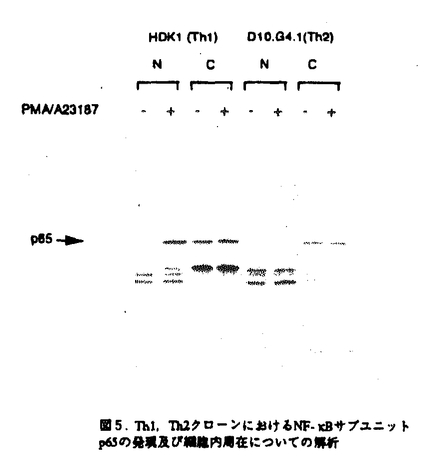
マウスのヘルパーT細胞は、そのサイトカイン産生パターンにより、Th1とTh2の2つのサブセットに分類され、生体内におけるこの2種類のヘルパーT細胞のバランスが免疫調節に重要であることが報告されている。しかし、このように異なるサイトカイン産生パターンがどのような機序で実現されるのかについては不明であった。また、cAMPは、Th1細胞とTh2細胞に対して選択的な作用を及ぼすこと、さらにTh2細胞内のcMAPレベルはTh1細胞のものより高いことが知られている。本論文では、Th1、Th2の相違の分子機構について明かにするために、IL-2遺伝子がTh1特異的に発現する機構及びcAMPによるIL-2遺伝子発現制御機構について解析を行っている。 本論文は序章、結果、考察からなる。結果は二部、考察は三部に分かれている。序章では、T細胞の活性化、IL-2遺伝子の発現制御機構、マウスヘルパーT細胞サブセット及びcAMPがT細胞に与える効果について概説している。 結果の第一部では、cAMPによるIL-2遺伝子発現制御機構について検討し、以下の新たな知見を得ている。(1)cAMPはIL-2遺伝子の転写開始点上流約300bpの領域を介してIL-2の転写を抑制する。 (2)NF-AT配列を介する転写がcAMPにより抑制される。(3)NF-ATの強制発現により、cAMPによるIL-2プロモーター活性の抑制はほぼ完全に回避される。(4)NF-AT配列結合複合体のAP-1構成因子がcAMPにより部分的にc-JunからJunBに移行する。 (5)NF-kB配列に対するp50/p65複合体の結合がcAMPにより抑制される。 これらの知見から、考察第一部において以下のように議論している。cAMPは、おそらくNF-AT複合体のNF-AT構成因子あるいはAP-1構成因子に何らかの作用を及ぼすことにより、NF-AT複合体の転写活性化能を抑制している。その機構の1つの可能性として、AP-1構成因子のc-JunからJunBへの移行が考えられる。また、NF-ATの強制発現によりcAMPの効果が回避されることから、cAMPはNF-AT複合体のAP-1構成因子のみならず、NF-AT構成因子にも何らかの作用を及ぼしていることが推察される。さらに、NF-kB配列に対するp50/p65複合体の結合がcAMPにより抑制されるという事実も考慮して、cAMPは、NF-AT、NF-kB、AP-1など複数の転写因子に対する作用を通してIL-2遺伝子発現を抑制すると結論づけられる。 結果の第二部では、IL-2遺伝子がTh1特異的に発現する機構について解析を行い、以下の事実を明かにしている。(1)IL-2のTh1特異的産生は、IL-2遺伝子の約300bpのプロモーターを介する転写レベルでの制御による。(2)NF-kB配列を介する転写誘導、NF-kB配列に対するNF-kBp50/p65複合体の結合誘導がTh1特異性を示す。(3)NF-kBp50/p65複合体の核移行はTh1細胞のみでPMAにより誘導される。(4)NF-kBの核移行の制御分子であるIkB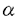 の分解の制御がTh1、Th2細胞で異なっている。 の分解の制御がTh1、Th2細胞で異なっている。 上記の事実から、考察第二部において以下のように議論している。NF-kB配列を介する転写誘導及びNF-kB配列に対するNF-kBp50/p65複合体のPMAによる結合誘導がTh1特異的であることから、NF-kBはIL-2プロモーター活性化のTh1特異性に寄与していることが示唆される。この現象は、Th1、Th2細胞においてIkBの分解の制御が異なっており、その結果としてNF-kBp50/p65複合体の核移行の制御が異なってくるために生じると考えられる。 最後に、考察第三部では、結果第一部及び第二部で得られた知見を総合することにより、IL-2遺伝子のみならず種々のサイトカイン遺伝子のcAMPによる制御機構及びThサブセット特異的制御機構を説明することを試みている。また、cAMPが引き起こす現象とTh2細胞でみられる現象が非常に類似しており、cAMPの作用機構とTh1、Th2の相違の分子機構が関連しているという仮説が支持されることについても指摘している。 以上の研究は、Th1、Th2細胞のサイトカイン産生パターンの相違の分子機構の解明に大きく貢献することが期待される。よって、論文提出者は、博士(理学)の学位を受けるのに十分な資格があるものと判断した。なお、本論文は、Hyun-Jun Lee博士、Esteban Sanai Masuda博士、小谷野-中川菜峰子博士、新井直子博士、新井賢-博士、横田崇博士との共同研究であるが、論文提出者が主体となって分析及び検証を行ったもので、論文提出者の寄与が十分であると判断した。 |  を産生し細胞性免疫に、Th2細胞はIL-4、IL-5を産生し液性免疫に関与する。Th1、Th2細胞が異なるサイトカイン産生パターンを示す機構については不明であるが、両細胞において、サイトカイン遺伝子発現に関与するシグナル伝達分子あるいは転写因子の制御が異なる可能性が示唆されている。また、マクロファージにより分泌されるプロスタグランジンE2などにより引き起こされる細胞内cAMPレベルの上昇は、Th1細胞とTh2細胞に対して相反する作用を及ぼすことが知られている。すなわち、cAMPレベルの上昇によりTh1細胞からのIL-2及びIFN
を産生し細胞性免疫に、Th2細胞はIL-4、IL-5を産生し液性免疫に関与する。Th1、Th2細胞が異なるサイトカイン産生パターンを示す機構については不明であるが、両細胞において、サイトカイン遺伝子発現に関与するシグナル伝達分子あるいは転写因子の制御が異なる可能性が示唆されている。また、マクロファージにより分泌されるプロスタグランジンE2などにより引き起こされる細胞内cAMPレベルの上昇は、Th1細胞とTh2細胞に対して相反する作用を及ぼすことが知られている。すなわち、cAMPレベルの上昇によりTh1細胞からのIL-2及びIFN