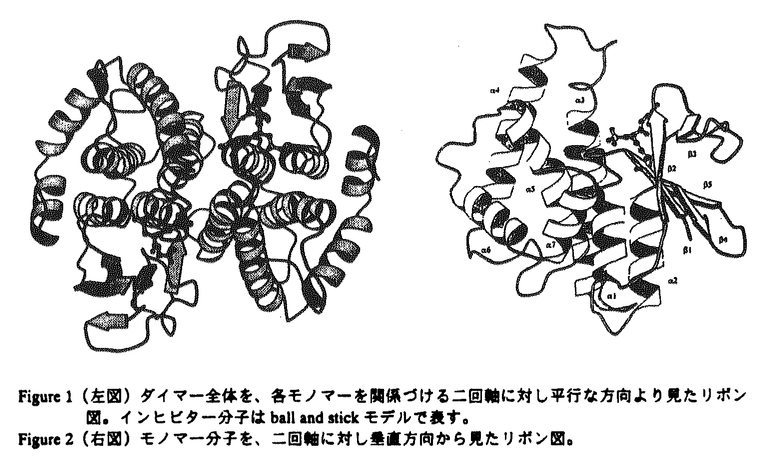
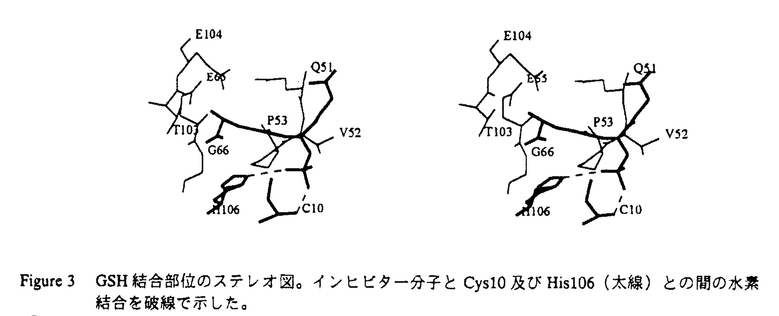
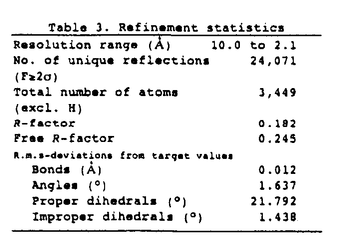
基本構造が同一である蛋白質分子間では、通常それらの触媒部位は、生物種を越えて高度に保存されている事が認められている。本論文では、原核生物由来のグルタチオンS-トランスフェラーゼ(GST)のグルタチオン(GSH)抱合反応の触媒機構は、真核生物由来のGSTファミリーと異なり、こうした一般論が成り立たない事を明らかにしている。 GSTは、解毒代謝に機能する多機能蛋白質であり、主としてグルタチオン(GSH)と求電子性基質との抱合反応を触媒する事により、これらの基質を無毒化する。GSTは、真核生物では幅広く存在する事が知られているが、他方原核生物では、その存在、性状、構造は全く不明であった。本論文では、既知のGSTとは最も遠縁の原核生物よりGSTを単離し、その三次元構造解析を行って、GSTファミリーの構造・機能の分子進化について知見を得る事を目的としている。 本論文では、2章1節で大腸菌よりGSTを単離、精製し、2節でそのクローニング及び各種性状解析を行っている。又、その大量発現系を構築し、部位特異的変異法による反応速度論的解析の結果を報告している。3節では、大腸菌由来GSTの結晶化、重原子同型置換法によるX線構造解析を行った。3章でその三次元構造を詳述し、4章で本酵素の触媒反応機構、及びその真核生物由来の酵素との差異に基づいてGSTファミリーの構造・機能相関の分子進化について考察を展開している。 2章1節では、まず、大腸菌よりGST活性を検索し、その高純度精製を行っている。これより本酵素のN端アミノ酸配列が明らかにされた。2節ではそのクローニングを行い、GST遺伝子の染色体上の位置、及びその構造遺伝子と周辺の塩基配列を同定した。次いで、その大量発現系を構築し、各種の酵素学的性状解析を行っている。本論文で、初めて原核生物由来のGSTの遺伝子の単離が行われたが、その真核生物由来のGSTに対する一次構造上の相同性は18%以下である。しかし、これらの実験結果から、大腸菌由来の酵素が分子量46kDaのホモ二量体であり、又、本酵素が、各種の基質に対するGSH抱合反応及びGSH依存的に過酸化物の還元反応を触媒する事から、既知のGSTファミリーに属する蛋白質である事が示唆された。 従来行われた多数の部位特異的変異による解析及びX線結晶構造解析の知見から、GSTファミリーでは、N端近傍に保存されるチロシン残基が活性中心であり、このアミノ酸残基がGSHのチオール基と直接相互作用してその求核性を高める事により、GSH抱合反応が触媒されると推測されている。ところが、本論文では、部位特異的変異法による反応速度度論的解析から、大腸菌由来の酵素ではN端近傍のTyr5が酵素活性に関与しない事を示している。これは、GSTファミリーにおいて、触媒部位に可変性が存在する事を示唆した初めての発見である。この問題を解明する為に、本論文では、更に2章3節で大腸菌由来GSTのX線結晶構造解析に研究を展開している。始めに大腸菌由来GSTの、GSH類似阻害剤グルタチオンスルフォネートとの複合体の結晶を得、これより重原子同型置換法により2.1Å分解能でのR因子18.2%の構造モデルを精密化した。この結果から、大腸菌由来GSTのフォールディングトポロジーは、基本的に既知のGSTと同一であり、特にGSH結合部位の構造とこれを構成するアミノ酸残基はよく保存されている事が明らかにされた。ところが、その触媒部位については、His106の側鎖及びCys10の主鎖のアミド基が阻害剤と近接した位置にあり、そのスルフォネート基と水素結合を形成している事が認められた。又、活性中心と予想されたチロシン残基は触媒部位に存在せず、2章2節における部位特異的変異の実験結果と符号する。これらの知見に基づいて、4章では、大腸菌由来GSTではCys10とHis106がチロシン残基に代わってGSHの求核性を高める事により、その抱合反応が触媒されると主張している。 以上の研究によって、GSTファミリーでは、原核生物由来と真核生物由来の酵素との間には、その触媒部位の構造と触媒機構に重大な相違が存在する事が示された。こうした例は酵素分子において、極めて希である事から、本研究より得られた知見は蛋白質の構造・機能相関の分子進化を研究する上において高い意義を持つと判断される。 又、本論文では、4章では原核生物由来と真核生物由来のGSTとの立体構造上の相違に基づいて、その生理機能の違いについて考察を展開している。例えば、そのC端領域の違いから、GSH抱合反応を受ける求電子性基質及び基質の認識に原核生物と真核生物由来のGSTでは違いのある可能性を推測している。GSTは、解毒代謝以外に、生理活性物質の生合成などにも機能しており、こうした基質認識の違いは、生体の代謝形成の進化を考察する上で貴重な知見といえる。 以上の、原核生物由来のGSTの検索、精製とそのクローニング、各種性状解析、部位特異的変異法による反応速度論的解析からX線結晶構造解析まで、これらはすべて、本論文提出者が主体となって行ったものであり、審査委員会は、本論文提出者が博士(理学)の学位をうける資格があるものと判定した。なお、本論文は、東京大学理学系研究科の横山茂之教授、同大学薬学部の佐藤能雅教授、東京薬科大学生命科学部の高橋健治教授との共同研究であるが、論文提出者が主体となって分析及び検証を行ったもので、論文提出者の寄与が十分であると判断した。 | 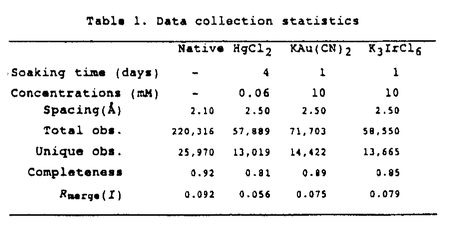
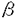 1〜
1〜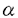 2)は2枚の
2)は2枚の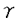 -glutamyl基を蛋白質分子内部に、glycyl基を外部に配向して結合しており、特に
-glutamyl基を蛋白質分子内部に、glycyl基を外部に配向して結合しており、特に 原子と水素結合を形成している(Figure3)。
原子と水素結合を形成している(Figure3)。